укороченный и ограниченный в росте репродуктивный побег, выполняющий функции микро- и мегаспорогенеза, опыления (осуществляемого при содействии агентов переноса пыльцы), оплодотворения, развития зародыша и образования плода. Ц. заканчивает собой стебель (главный или боковой). Часть стебля под Ц., обычно лишённая листьев, называется цветоножкой. Нередко она очень укорочена и едва выражена, и тогда Ц. называется сидячим. Цветоножка переходит в укороченную ось Ц. - цветоложе, которое обычно шире цветоножки и характеризуется слабо выраженными междоузлиями. Цветоложе может быть различной формы. На его узлах расположены все части цветка, как стерильные (чашелистики и лепестки), так и фертильные (тычинки и плодолистики). Чашелистики, образующие в совокупности чашечку, обычно служат органами защиты Ц., особенно до его распускания (в бутоне), а часто также дополнительными органами фотосинтеза. Нередко они становятся лепестковидными и принимают на себя функцию лепестков, например у ломоноса. В процессе эволюции Ц. чашелистики часто срастаются в сростнолистную чашечку. Лепестки служат для привлечения опылителей. Совокупность лепестков Ц. называется венчиком. Чашечка и венчик составляют вместе околоцветник, или покров цветка. Когда в цветке имеются как чашечка, так и венчик, то околоцветник называется двойным. В тех случаях, когда лепестки отсутствуют или когда нет ясно выраженных различий между чашечкой и венчиком, околоцветник называется простым. В центре Ц. расположены тычинки и плодолистики. Как по своей функции, так и по происхождению тычинки представляют собой
Микроспорофиллы
. Наиболее примитивные из них (у ряда представителей семейства магнолиевых и винтеровых) представляют собой широкую лентовидную пластинку, снабженную четырьмя попарно сближенными линейными микроспорангиями, более или менее погруженными в ткань пластинки. В процессе эволюции тычинки постепенно дифференцировались на тычиночную нить и пыльник, состоящий из двух половинок, соединённых связником (продолжение тычиночной нити). Совокупность тычинок в цветке называется андроцеем. Тычиночные нити часто срастаются как между собой, так и с членами околоцветника и с плодолистиками. Иногда тычинки срастаются между собой также пыльниками (нередко только пыльниками). Плодолистики, или карпеллы, представляют собой сильно видоизменённые и специализированные
Мегаспорофиллы
. Нередко плодолистик называется пестиком (простым пестиком), однако этот термин применяется в разных смыслах и вносит путаницу. Наиболее примитивный тип плодолистика сохранился у родов дегенерия (семейство дегенериевые) и рода Тасмания из семейства винтеровых. Он состоит из короткой ножки (гиноподия) и относительно тонкой адаксиально сложенной (кондупликатной) пластинки и внешне очень напоминает сложенные в почки кондупликатные листья. Вдоль плодолистика проходят 3 проводящих пучка (жилки), из которых средняя, или спинная (дорсальная), сильно ветвится. Между средней и боковыми жилками расположены многочисленные семязачатки, или семяпочки. На ранних стадиях развития края плодолистиков дегенерии почти не соприкасаются между собой, а защита семязачатков во время цветения обеспечивается благодаря тесному сближению широких боковых участков плодолистика, расположенных между его краями и плацентами. Срастание этих сближенных внутренних поверхностей происходит лишь после цветения, т. е. в процессе развития плода. У др. представителей винтеровых наблюдаются самые различные стадии срастания. У всех цветковых растений на плодолистике имеется особая, служащая для улавливания пыльцы железистая ткань, называемая рыльцем. У дегенерии и Тасмании рыльце примитивное и протягивается по всей длине плодолистика. Верхняя часть плодолистика, в которой оно локализуется, обычно вытягивается в тонкий стерильный столбиковидный вырост, приподнимающий рыльце над фертильной частью (завязью) плодолистика и служащий для прохождения пыльцевой трубки. Эта специализированная тонкая часть плодолистика называется столбиком (нередко её называют стилодием, оставляя термин столбик только для сросшихся стилодиев). На ранних стадиях эволюции столбика (стилодия) он характеризуется низбегающим рыльцем, но в процессе дальнейшей эволюции рыльце постепенно локализуется на верхушке (головчатое рыльце). Совокупность плодолистиков в цветке называется гинецеем. Наиболее примитивные типы гинецея со свободными плодолистиками (например, у магнолии, лютика, пиона) называются апокарпными. В процессе эволюции плодолистики постепенно срастаются между собой, и из апокарпного гинецея возникает ценокарпный (см.
Ценокарпия)
, в котором стилодии могут оставаться свободными или же срастаться, образуя столбик. Эволюция ценокарпного гинецея, как и эволюция апокарпного, обычно сопровождается уменьшением числа плодолистиков и в большинстве случаев также числа семязачатков. Крайняя форма редукции числа семязачатков в ценокарпном гинецее - т. н. псевдомономерный гинецей, у которого фертилен и полностью развит один плодолистик (у ильма, шелковицы, крапивы, конопли и др.).
В Ц. большинства цветковых растений имеются специальные экскреторные органы для производства нектара - нектарники. Они возникли в самых разных ветвях эволюции цветковых растений и на самой разной морфологической основе, т. е. на самых разных частях Ц.
Примитивные типы Ц. характеризуются относительно длинным цветоложем, но в некоторых случаях, как, например, у мышехвостика или у некоторых магнолиевых, имеется сильно удлинённое цветоложе вторичного происхождения. В процессе эволюции происходит постепенное укорочение цветоложа, в связи с чем части Ц. всё более сближаются и первоначально спиральное их расположение изменяется на циклическое (круговое). Части циклического Ц. обычно расположены мутовчато, реже - супротивно. Они встречаются гораздо чаще спиральных. Промежуточное положение занимают спироциклические Ц., например Ц. лютика. Примитивные Ц. - актиноморфные ("правильные"), т. е. такие, которые могут быть разделены вертикальной плоскостью, проходящей через ось симметрии, на равные половины не менее чем в двух направлениях. В процессе эволюции (связанной с биологией опыления (См.
Опыление)) из актиноморфных возникли зигоморфные ("неправильные") Ц. Они могут быть разделены на две половины только в одном направлении (например,
цветок гороха или львиного зева). В отличие от симметричных, т. е. актиноморфных и зигоморфных Ц., встречаются также асимметричные Ц., которые невозможно разделить плоскостью симметрии на две равные половины, например у канны.
Ц. могут быть обоеполыми или однополыми (раздельнополыми). Однополые Ц. произошли из обоеполых в результате редукции в одних случаях - тычинок, в других - плодолистиков. В однополых Ц. часто сохраняются рудименты тычинок и плодолистиков. Предполагают, что Ц. произошёл из репродуктивных побегов гипотетических предков цветковых растений - семенных папоротников. В результате укорочения оси побега она превратилась в цветоложе, а в результате упрощения их расчленённых микро- и мегаспорофиллов возникли простые, неразветвлённые тычинки и плодолистики цветковых растений. [Предположение, что Ц. произошёл из стробила (шишки) саговников (цикадовых), гнетовых или беннеттитовых, маловероятно.] При этом эволюция Ц., как и эволюция стробилов разных групп голосеменных, пошла разными путями. Т. о., современные представления основаны на предположении, что основные части Ц. - тычинки и плодолистики - произошли из спороносных органов предков цветковых растений. Трудно сказать с определённостью, были ли эти спороношения трёхмерными или плоскими, лишёнными хлорофилла или зелёными. Вероятнее всего, они представляли собой плоские и более или менее упрощённые (возможно, лопастные) спорофиллы, которые (особенно мегаспорофиллы), вероятно, частично несли функцию фотосинтеза.
Лит., Кернер фон-Марилаун А., Жизнь растений, пер. с нем., т. 2, СПБ, 1900; Голенкин М. И., Курс высших растений, М. - Л., 1937; Имс А., Морфология цветковых растений, пер. с англ., М., 1964; Тахгаджян А. Т., Основы эволюционной морфологии покрытосеменных, М. - Л., 1964; Первухина Н. В., Проблемы морфологии и биологии цветка, Л., 1970; Эсау К., Анатомия растений, пер. с англ., М., 1969; Федоров А. А., Артюшенко З. Т., Атлас по описательной морфологии высших растений, [т. 3] - Цвет
ок, Л., 1975. См. также лит. к ст.
Цветковые растения.
А. Л. Тахтаджян.
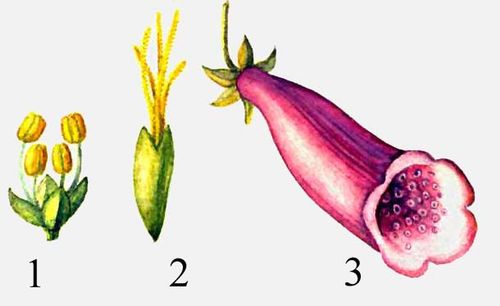
Цветки: 1 - лебеды (тычиночный цветок), 2 - лебеды (пестичный цветок), 3 - наперстянки.
Цветки: 1 - лилии, 2 - василька (воронковидный цветок), 3 - василька (трубчатый цветок), 4 - погремка, 5 - опунции, 6 - тюльпана, 7 - табака.
Цветки: 1 - кувшинки, 2 - цикория.
Цветки: 1 - розы, 2 - кирказона.
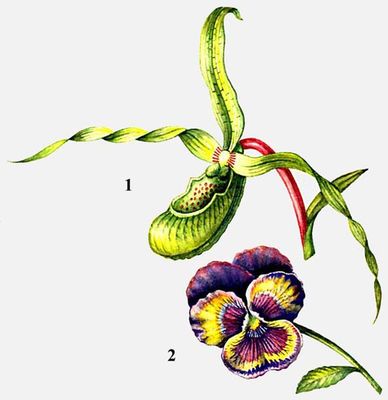
Цветки: 1 - соленипедиума, 2 - фиалки.
Цветки: 1 - нарциса, 2 -ясеня, 3 - пшеницы, 4 - вяза, 5 - камыша, 6 - страстоцвета.

![рудбекии блестящей]]](https://commons.wikimedia.org/wiki/Special:FilePath/Blume mit Schmetterling und Biene 1uf.JPG?width=200 "рудбекии блестящей]]")


![Цветок [[губастик]]а в видимом свете (слева) и в ультрафиолете (справа), тёмная область на правом снимке — указатель нектара](https://commons.wikimedia.org/wiki/Special:FilePath/Mimulus nectar guide UV VIS.jpg?width=200 "Цветок [[губастик]]а в видимом свете (слева) и в ультрафиолете (справа), тёмная область на правом снимке — указатель нектара")
![живокости]], с пятью голубыми чашелистиками и белым глазком, образованным лепестками-нектарниками и лепестками-стаминодиями](https://commons.wikimedia.org/wiki/Special:FilePath/Дельфиниум.JPG?width=200 "живокости]], с пятью голубыми чашелистиками и белым глазком, образованным лепестками-нектарниками и лепестками-стаминодиями")