человека (франц., единственное число race), исторически сложившиеся ареальные (см.
Ареал) группы людей, связанные единством происхождения, которое выражается в общих наследственных морфологических и физиологических признаках, варьирующих в определённых пределах. Т. к. групповая и индивидуальная изменчивость этих признаков не совпадает, Р. являются не совокупностями особей, а совокупностями популяций (См.
Популяция), т. е. территориальных групп людей, объединяемых брачными связями. Р. - внутривидовые таксономические (систематические) категории, находящиеся в состоянии динамического равновесия, т. е. изменяющиеся в пространстве и во времени во взаимодействии с окружающей средой и вместе с тем обладающие определённой, генетически обусловленной устойчивостью. По всем основным морфологическим, физиологическим и психологическим особенностям, характерным для современных людей, сходство между всеми Р. велико, а различия несущественны. Лишены всякого фактического основания реакционные концепции о существовании "высших" и "низших"
Р. (см.
Расизм) и об их происхождении от разных родов высших обезьян (см.
Полигенизм). Данные антропологии и др. наук доказывают, что все Р. происходят от одного вида ископаемых гоминид (см.
Моногенизм). Неограниченные возможности смешения (метисации (См.
Метисация)) между всеми Р., полная биологическая и социально-культурная полноценность смешанных групп служат веским доказательством видового единства человечества и несостоятельности расизма. Слово "Р.", восходящее, вероятно, к арабскому корню "
рас" (голова, начало), встречается впервые в современном смысле у французского учёного Ф. Бернье (1684). В 18-20 вв. были предложены многочисленные классификации Р., основанные главным образом на внешних морфологических особенностях. Одна из наиболее удачных классификаций принадлежит Ж.
Деникеру (1900).
Основные группы Р. Наиболее отчётливо в составе современного человечества выделяются три основные группы Р. - негроидная, европеоидная и монголоидная; их часто называют большими Р. Для негроидов характерны курчавые чёрные волосы, тёмно-коричневая кожа, карие глаза, слабое или среднее развитие третичного волосяного покрова, умеренное выступание скул, сильно выступающие челюсти (
Прогнатизм), слабо выступающий широкий нос, часто с поперечно, т. е. параллельно плоскости лица, расположенными ноздрями, утолщённые губы. Европеоиды отличаются волнистыми или прямыми мягкими волосами разных оттенков, сравнительно светлой кожей, большим разнообразием окраски радужины глаз (от карих до светло-серых и голубых), сильным развитием третичного волосяного покрова (в частности, бороды у мужчин), слабым выступанием скул, незначительным выступанием челюстей (
Ортогнатизм), узким выступающим носом с высоким переносьем, обычно тонкими или средними губами. Монголоидам свойственны прямые жёсткие тёмные волосы, слабое развитие третичного волосяного покрова, желтоватые оттенки кожи, карие глаза, уплощённое лицо с сильно выдающимися скулами, узкий или среднеширокий нос с низким переносьем, умеренно утолщённые губы, наличие особой кожной складки верхнего века, прикрывающей слёзный бугорок во внутренних углах глаз (см.
Эпикантус). К монголоидным Р. по происхождению и многим признакам близки американские индейцы (см.
Американская раса), у которых, однако, эпикантус встречается редко, нос выступает обычно сильно, общий монголоидный облик часто бывает сглажен.
Далеко не все популяции современного человечества могут быть отнесены к трём описанным основным группам Р. На Ю.-В. Азии, в Океании и Австралии широко расселены популяции, по некоторым признакам (тёмная окраска кожи, широкий нос, толстые губы) близкие к негроидам Африки, но отличающиеся от них волнистыми волосами, сильным развитием третичного волосяного покрова, иногда ослабленной пигментацией (См.
Пигментация)
. Эти популяции одни учёные рассматривают как четвёртую основную группу Р. - австралоидную, другие же объединяют с негроидами в большую экваториальную (негро-австралоидную) расу (См.
Экваториальная раса). У австралоидов очень велик свойственный всему человечеству
Генетический полиморфизм, который находит внешнее выражение в огромном групповом разнообразии сочетаний расовых признаков. Так, например, аборигены Австралии по пигментации близки к африканским негроидам, а по форме волос и развитию третичного волосяного покрова - к европеоидам (см.
Австралийская раса). Для папуасов и меланезийцев характерно сочетание многих австралоидных черт с курчавыми волосами (см.
Меланезийская раса). Для веддоидов, представленных веддами Шри-Ланки и др. малыми народами Южной и Юго-Восточной Азии, типична комбинация общего австралоидного облика с малым ростом, слабым развитием бороды и надбровных дуг. Генетически с австралоидными Р. связаны, по мнению многих учёных, также курчавоволосые крайне низкорослые негритосы, а также айны, у которых относительно светлая кожа и наиболее обильный в мире рост волос на лице и теле сочетаются с некоторыми монголоидными особенностями (уплощённость лица, эпикантус).
Негроидные особенности наиболее выражены в африканских популяциях, расселённых к Ю. от Сахары и известных под собирательным и неточным названием негры. Кроме того, к негроидам относятся крайне низкорослые центральноафриканские
Пигмеи, или негрилли, внешне сходные с азиатскими негритосами. С негроидами некоторые антропологи сближают также южноафриканских бушменов и готтентотов, у которых крайняя степень курчавости сочетается с отдельными монголоидными чертами (желтоватая кожа, уплощённое лицо, эпикантус). Между ареалами экваториальных (австралоидных и негроидных) Р. и расселённых севернее европеоидов расположен широкий пояс переходных групп, из которых одни, известные с глубокой древности, отражают генетические связи между обеими группами упомянутых Р., другие же сложились в средние века и в новое время в процессе межрасового смешения - метисации. К древним переходным группам относятся южноиндийская (дравидская) и восточноафриканская, или эфиопская, Р. Последняя по цвету кожи почти не отличается от негров, а по строению лица и форме носа напоминает юж. европеоидов. Промежуточность по большинству расовых признаков проявляется также во многих популяциях Судана, особенно у народа
Фульбе. Европеоидные Р., сформировавшиеся первоначально в Юго-Западной Азии, Северной Африке и Европе, могут быть подразделены на три главные группы: южную - со смуглой кожей, тёмными глазами и волосами; северную - со светлой кожей, значительной долей серых и голубых глаз, русых и белокурых волос; промежуточную, для которой характерна средне-интенсивная пигментация. По окраске кожи, глаз и волос, по строению лицевого скелета и мягких частей лица, по пропорциям мозговой части черепа, часто выражаемым головным указателем (См.
Головной указатель), и по некоторым др. признакам среди европеоидов антропологи выделяют различные локальные Р., или т. н. Р. второго порядка. Южных европеоидов в целом, учитывая их ареал, называют индо-средиземноморской расой (См.
Индо-средиземноморская раса)
. Среди относительно длинноголовых (см.
Долихокефалия)
популяций этой Р. выделяют собственно средиземноморскую (медитерранную) на З. и индо-афганскую на В. В составе короткоголовых (см.
Брахикефалия) южных европеоидов - адриатическую, или динарскую расу (См.
Динарская раса), переднеазиатскую расу (арпеноидную) и памиро-ферганскую расу (См.
Памиро-Ферганская раса)
.
Промежуточных по пигментации европеоидов, большей частью короткоголовых, подразделяют на альпийскую, среднеевропейскую и др. Р. Мезодолихокефальных (среднедлинноголовых) светлых европеоидов раньше описывали под названием северной, или нордийской, Р., а более брахикефальных - под названием балтийской Р. Некоторые антропологи всех светлых европеоидов подразделяют на северо-западных (см.
Атланто-балтийская раса) и северо-восточных (см.
Беломорско-балтийская раса); в формировании последних могли принимать участие древние монголоидные популяции, проникавшие в Европу из-за Урала.
На восточных рубежах своего ареала европеоиды с древнейших времён взаимодействовали с монголоидами. В результате их раннего смешения, начавшегося, вероятно, ещё в эпоху
Мезолита, сложилась на С.-З. Сибири и на крайнем В. Европы
Уральская раса, для которой характерно сочетание промежуточных монголоидно-европеоидных особенностей с некоторыми специфическими чертами (например, с вогнутой формой спинки носа). К уральской Р. по многим признакам близка
Лапоноидная раса; многие антропологи даже объединяют обе эти Р. в одну (урало-лапоноидную). Позднее (с первых веков н. э.) в степной полосе между Уралом и Енисеем формируется в процессе смешения монголоидов и европеоидов южно-сибирская раса (См.
Южносибирская раса) с очень широким лицом и выраженной брахикефалией. В средние века на территории Средней Азии складываются новые смешанные европеоидно-монголоидные популяции. Собственно монголоидные Р. в Азии подразделяются па две главные группы - континентальную и тихоокеанскую; первая отличается от второй более светлой кожей, некоторой тенденцией к депигментации волос и глаз, очень крупными размерами лица,
Ортогнатизмом,
более тонкими губами. В составе континентальных монголоидов выделяются сибирская, или североазиатская, и центральноазиатская Р. Промежуточное положение между континентальными и тихоокеанскими монголоидами занимает арктическая (эскимосская) Р. с крайне высоким и широким лицом, тенденцией к прогнатизму и очень узким носом. Северные группы тихоокеанских монголоидов с высоким, но сравнительно узким лицом объединяются в дальневосточную, или восточноазиатскую, Р. Классификация американских монголоидов, происходящих, несомненно, из Азии, очень затруднена, т.к. многие группы индейцев были истреблены колонизаторами, оттеснены с первоначальных мест расселения или смешались с европейцами или африканцами.
Южные группы тихоокеанских монголоидов, входящие в состав южноазиатской, или малайской
расы (См.
Малайская раса), обнаруживают немало австралоидных особенностей: наличие волнистых волос, иногда довольно обильный рост бороды и волос на теле, тёмная кожа оливковых оттенков, низкое лицо, относительно широкий нос, утолщённые губы и др. Многие из перечисленных особенностей свойственны японцам, в формировании расового состава которых приняли участие монголоидные и австралоидные компоненты (последние, вероятно, были частично связаны с айнами). На В. Индонезии интенсивная метисация тихоокеанских монголоидов с папуасами также привела к формированию промежуточных популяций. Во многом аналогичный процесс имел место и на Мадагаскаре, куда, по-видимому, уже в 1-м тыс. до н. э. из Индонезии переселились различные южноазиатские группы, смешавшиеся на острове с негроидами. Очень своеобразные сочетания монголоидных, австралоидных, а иногда и европеоидных черт характерны для микронезийцев (См.
Микронезийцы) и особенно для полинезийцев (См.
Полинезийцы)
. Почти все упомянутые выше расовые признаки наследуются независимо друг от друга и являются полигенными, т. е. контролируемыми многими
Генами
. Но у людей существуют и др. ареальные особенности с более простой генетической структурой, зависящие от одной или немногих пар аллелей (См.
Аллели)
. К ним принадлежат многие эритроцитарные группы крови, белки сыворотки, некоторые детали строения зубов (см.
Одонтология), узоры на подушечках пальцев рук и ног (см.
Дерматоглифика), вкусовые ощущения при пробе на фенилтиокарбамид, виды цветовой слепоты (см.
Дальтонизм) и многие др. морфофизиологические и биохимические особенности, географические вариации которых не вполне совпадают с ареалами основных Р., хотя и обнаруживают в пределах каждой из них определённые закономерности распределения. Анализ изменчивости всей совокупности расовых признаков позволяет поставить вопрос о разделении Р. по их генетическим связям на две группы - западную и восточную. Одни учёные (например, советский антрополог В. П. Алексеев), основываясь главным образом на особенностях волосяного покрова и строения черепа, относят к западной группе европеоидную и экваториальную (негро-австралоидную) большие Р., а к восточной - монголоидную. Др. исследователи (например, советские антропологи А. А. Зубов, Н. Н. Чебоксаров), используя данные одонтологии, дерматоглифики и серологии, в западную (атланто-средиземноморскую) группу Р. включают негроидов и европеоидов, а в восточную (тихоокеанскую) - австралоидов и монголоидов; вторая группа отличается от первой большей долей резцов лопатообразной формы и др. деталями строения зубов, высокой частотой круговых узоров на подушечках пальцев, специфическим распределением
Генотипов и
Фенотипов по многим серологическим системам (например, почти 100\%-ной концентрацией резус-положительности).
История современных Р. Одни антропологи предполагают, что Р. начали складываться у древнейших людей, (архантропов (См.
Архантропы)) в нескольких центрах Африки, Европы и Азии (см.
Полицентризм), другие же (в т. ч. большинство советских учёных) считают, что расовая дифференциация происходила позднее, уже после образования человека современного вида в Восточном Средиземноморье и соседних областях Южной Европы, Северной и Восточной Африки и Западной Азии (см.
Моноцентризм). Вероятно, первоначально в конце палеолита у людей современного вида возникли два очага расообразования: западный - на С.-В. Африки и на Ю.-З. Азии и восточный - на В. и Ю.-В. Азии. Позднее различные популяции людей, расселяясь по земному шару, смешивались между собой и, приспосабливаясь к различным естественно-географическим условиям, распадались на современные Р.
Многие расовые признаки, возникшие первоначально путём мутаций (См.
Мутация), приобрели приспособительное значение и под действием естественного отбора на ранних этапах расогенеза закреплялись и распространялись в популяциях, живших в разной географической среде. Характерные особенности негроидных и австралоидных Р. складывались в Африке и Южной Азии в условиях жаркого влажного климата с усиленным солнечным освещением, от вредного действия которого могла предохранять тёмная окраска кожи, а возможно, и курчавые волосы, образующие на голове естественную защитную "шапку". В тропиках приспособительное значение для усиленного испарения влаги через слизистую оболочку могли иметь утолщённые губы и поперечно расположенные широко открытые ноздри. У европеоидов действию отбора могла подвергаться светлая окраска кожи, волос, глаз, т.к. мутации, определяющие эти признаки, имели наибольшие шансы выживания и распространения в Северной и Средней Европе, где в позднем палеолите преобладал прохладный влажный климат со значительной облачностью и пониженной инсоляцией. У монголоидных Р., формировавшихся, вероятно, в степях и полупустынях Центральной Азии, приспособительную роль могли играть эпикаптус и сильно развитая складка верхнего века, защищавшие глаза от сильных ветров и песчаных бурь, очень характерных для сухого континентального климата с резкими суточными и сезонными колебаниями температуры. Отбор мог играть известную роль в распределении некоторых серологических признаков, например в повышенной концентрации группы В системы АВО (см.
Группы крови) в Китае, Индии и др. странах Азии, где были часты эпидемии оспы (люди с этой группой реже заболевают оспой и легче её переносят).
С развитием производительных сил общества и созданием в процессе коллективного труда искусственной культурной среды роль естественного отбора в расогенезе постепенно уменьшалась. Снижение это началось ещё, вероятно, при переходе от палеолита к мезолиту, когда люди из Северо-Восточной Азии стали заселять Америку, а из Юго-Восточной Азии - Австралию и крупные острова Океании. Приспособительный характер расовых особенностей населения этих стран выражен слабее, чем у человеческих групп заселённых ранее районов Африки, Азии и Европы. Значительную роль в расогенезе играла изоляция отдельных, особенно малых популяций, в которых при заключении на протяжении многих поколений браков преимущественно внутри своей группы (см.
Эндогамия) могли происходить заметные сдвиги в распределении
Генов,
контролирующих расовые признаки. Процессы эти известны в генетике под названием генетико-автоматических процессов (См.
Генетико-автоматические процессы), или дрейфа генов; ими легче всего объяснить возникновение многих одонтологических, серологических, дерматоглифических и др., большей частью нейтральных, различий между разными популяциями (в частности, между западными и восточными группами основных Р.). При заселении Америки генетико-автоматические процессы привели к почти полному исчезновению группы В и к сильному снижению доли группы А системы АВО у индейцев. У австралийских аборигенов дрейф генов, напротив, вызвал увеличение частоты группы А. Если роль отбора и изоляции в расогенезе непрерывно падала, то роль метисации, напротив, увеличивалась по мере роста связей между разными странами, переселений и миграций. В 16-19 вв. в результате вольных и невольных переселений европейцев и африканцев возникли новые метисные группы: американские
Мулаты и
Метисы, южно-африканские "цветные", смешанные группы населения Сибири и др. Конкретная история расового состава народов, изучаемая этнической антропологией (См.
Этническая антропология), зависит от характера их взаимодействия, колебаний численности, от типа хозяйства и культурного уровня населения, от интенсивности эксплуатации угнетённых классов, от образования и разрушения генетических барьеров (географических, сословно-кастовых, профессиональных и др.), от таких явлений, как войны, голод, эпидемии и т.п., от процессов этнической ассимиляции (См.
Ассимиляция) и интеграции (см.
Этногенез,
Этнические процессы)
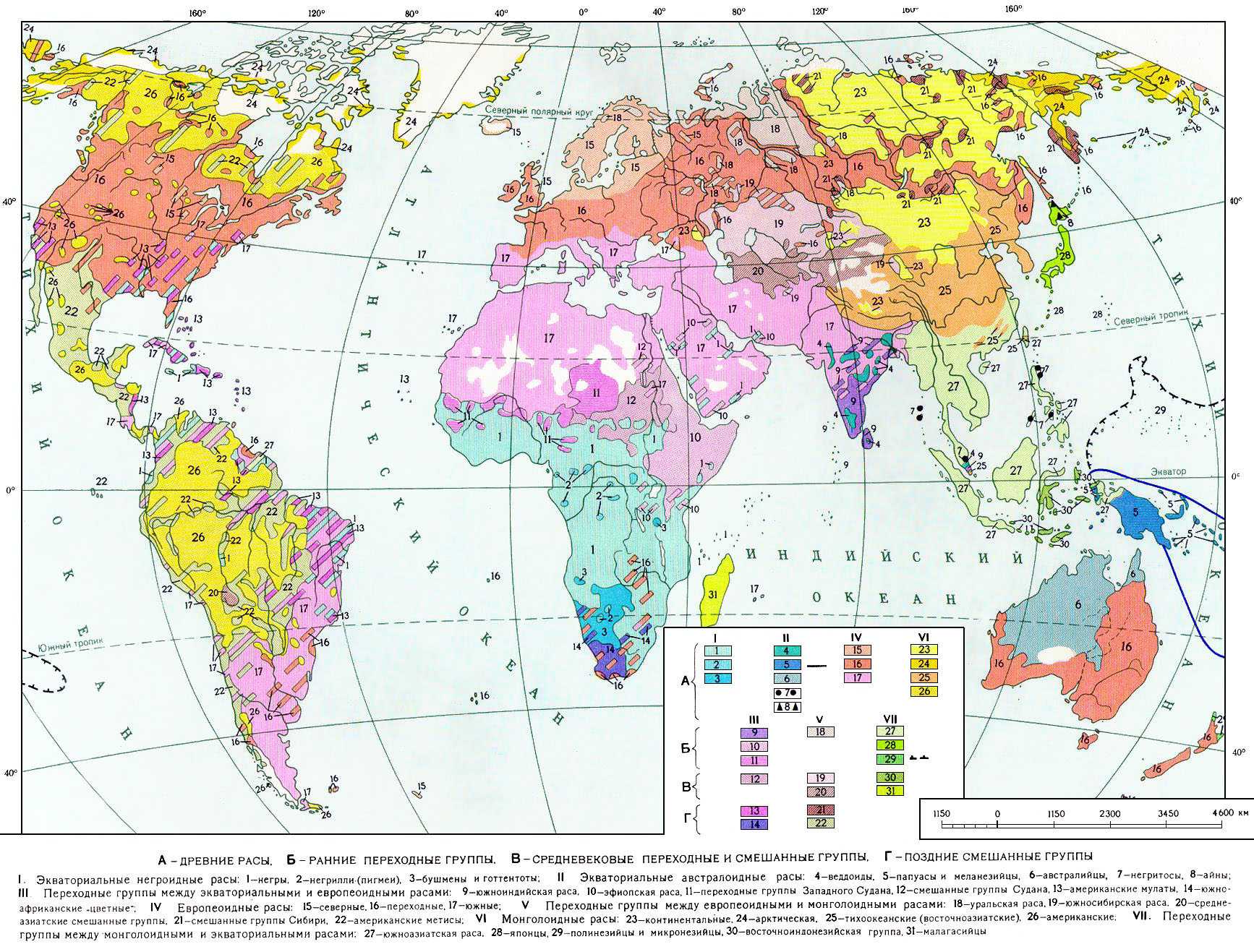
. По мере развития экономического, социального и культурного, а также биологического взаимодействия между различными народами, границы расовых ареалов всё больше и больше стираются, возникают новые местные сочетания различных расовых признаков единого человечества. (См. карту.)
Лит.: Наука о расах и расизм, М., 1939; Дебец Г. Ф., Палеоантропология СССР, М. -Л., 1948; Рогинский Я. Я., Что такое человеческие расы, М., 1948; Происхождение человека и древнее расселение человечества, М., 1951; Бунак В. В., Человеческие расы и пути их образования, "Советская этнография", 1956, № 1; Рогинский Я. Я., Левин М. Г., Антропология, М., 1963; Дебец Г. Ф., Расовый состав мира, в книга: Атлас народов мира, М., 1964; Нестурх М. Ф., Человеческие расы, М., 1965; Рогинский Я. Я., Рынков Ю. Г., Генетика расообразования у человека, в книге: Проблемы медицинской генетики, М., 1970; Чебоксаров Н. Н., Чебоксарова И. А., Народы, расы, культуры, М., 1971: их же, Этносы, популяции, расы, в сборнике: Земля и люди, М., 1974: Расы и народы. Ежегодник, в. 1-3, М., 1971-73; Арутюнов С. А., Чебоксаров Н. Н., Этнические процессы и информация, "Природа", 1972, № 7; Алексеев В. П., В поисках предков. Антропология и история, М., 1972; его же, География человеческих рас, М., 1974; Бромлей Ю. В., Этнос и этнография, М., 1973; Зубов А. А., Этническая одонтология, М., 1973; Montagu Ashley (ed.), The concept of race, L., 1969; Coon L. S., Hunt Е. Е., The living races of man, N. Y., 1965; Schwidetzky 1., Die neue Rassenkunde, Stuttg., 1962.
Н. Н. Чебоксаров.

![Иоганна Блуменбаха]]. Издание 1795 года](https://commons.wikimedia.org/wiki/Special:FilePath/Blumenbach's five races.JPG?width=200 "Иоганна Блуменбаха]]. Издание 1795 года")

![Кавказа]]](https://commons.wikimedia.org/wiki/Special:FilePath/Georgierin.png?width=200 "Кавказа]]")
![Густав Мютцель]], 1893, Универсальная история Ридпата, 1897](https://commons.wikimedia.org/wiki/Special:FilePath/Gustav Mützel - Principal types of mankind (after Huxley), 1893.jpg?width=200 "Густав Мютцель]], 1893, Универсальная история Ридпата, 1897")
![американская, или медного цвета]]» и «малайская, или оливкового цвета»](https://commons.wikimedia.org/wiki/Special:FilePath/Mankind1ma6.jpg?width=200 "американская, или медного цвета]]» и «малайская, или оливкового цвета»")

![трёх рас]]](https://commons.wikimedia.org/wiki/Special:FilePath/Stamp of USSR 2022.jpg?width=200 "трёх рас]]")

![митохондриальных]] [[ДНК]] (в тысячах лет до настоящего времени)](https://commons.wikimedia.org/wiki/Special:FilePath/World map of prehistoric human migrations.jpg?width=200 "митохондриальных]] [[ДНК]] (в тысячах лет до настоящего времени)")