Voer een woord of zin in in een taal naar keuze 👆
Taal:
Vertaling en analyse van woorden door kunstmatige intelligentie ChatGPT
Op deze pagina kunt u een gedetailleerde analyse krijgen van een woord of zin, geproduceerd met behulp van de beste kunstmatige intelligentietechnologie tot nu toe:
- hoe het woord wordt gebruikt
- gebruiksfrequentie
- het wordt vaker gebruikt in mondelinge of schriftelijke toespraken
- opties voor woordvertaling
- Gebruiksvoorbeelden (meerdere zinnen met vertaling)
- etymologie
Wat (wie) is Водяной побег - definitie
СТАТЬЯ-СПИСОК В ПРОЕКТЕ ВИКИМЕДИА
Побег (сезон 3); Побег (сезон 2); Побег (сезон 4); Побег (сезон 5); Побег (сезон 1)

Водяной побег
волчок, жировой побег, побег на стволе или толстой ветви дерева, развившийся из "спящей" почки. Имеет более крупные, чем на других побегах, листья. В. п. образуются при обмерзании, обрезке, увеличении освещения дерева (например, при вырубке соседних деревьев) и т.п.; наблюдаются часто у дубов, клёнов, вязов, осокорей, пирамидальных тополей и др. На плодовых деревьях В. п. обычно уничтожают, так как развитие их уменьшает число цветочных почек, а значит и плодов.
Побег
СТРАНИЦА ЗНАЧЕНИЙ В ПРОЕКТЕ ВИКИМЕДИА
Побег (фильм); Побег (фильм, 2005)
I
Побе́г (cormus)
в ботанике, один из основных органов высших растений; состоит из стебля (См. Стебель) и сидящих на нём Листьев и почек (См. Почка). Система П. вместе с корневой системой составляет тело папоротников, хвощей, плаунов, голосеменных и покрытосеменных растений, что послужило основанием для особого их наименования - Кормофиты, или побегоносные растения (в отличие от таллофитов (См. Таллофиты)). П. у высших растений возникли в процессе филогенеза как приспособление к наземному образу жизни. У первенцев наземной флоры - псилофитов (см. Риниофиты) - П. и корней ещё не было; их дихотомически ветвящееся тело состояло из вегетативных и спороносных безлистных веточек - теломов. П. сформировались в результате агрегации ("скучивания") и слияния теломов, причём листья возникли либо как поверхностные выросты на осях (микрофильная линия эволюции - плауны), либо вследствие уплощения и срастания целых систем теломов (макрофильная линия - папоротникообразные и семенные растения). Общность происхождения, единство проводящей системы листьев и стеблей и формирование их в онтогенезе из единой массы меристемы (См. Меристема) (конуса нарастания) подчёркивают целостность П. как единого органа.
Возникновение П., т. е. листостебельности, - крупнейший этап в развитии растительного мира на Земле. Благодаря плоской форме листьев резко возросла фотосинтезирующая поверхность; связанное с этим усиление транспирации (См. Транспирация) способствовало развитию настоящих корней как совершенных органов поглощения воды и минеральных солей. В результате побегоносные растения заняли всю поверхность суши Земли и начиная с каменноугольного периода стали господствовать в её растительном покрове.
Первичные формы П. - ассимилирующие и спороносные - выполняли лишь основные функции: Фотосинтеза и размножения. Впоследствии функции П. стали разнообразнее. Структура типичного ассимилирующего П. отражает разделение важнейших жизненных функций между его отдельными органами. Листья - органы фотосинтеза и транспирации. Стебли - опорные органы, на которых листья располагаются в наиболее выгодном для фотосинтеза положении (см. Листорасположение), для проведения воды, солей и пластических веществ. Почки - органы нарастания, возобновления и вегетативного размножения. В них заключены очаги меристемы, способные обеспечивать верхушечный рост П. (верхушечная почка) и его Ветвление, т. е. образование П. следующих порядков, формирование системы П. (боковые почки). Для структуры П. характерна метамерность, т. е. повторяемость строения его частей по продольной оси. Структурные элементы П. - узел с отходящим от него листом или мутовкой листьев и междоузлие. Почки обычно располагаются в пазухах листьев. Метамеры, последовательно появляющиеся на конусе нарастания П., закономерно изменяются от его основания до верхушки.
У однолетних растений все П. живут только один сезон. У многолетних - длительность жизни П. может быть разной, но ежегодно из почек появляются П. возобновления, входящие в состав многолетней системы П. и заменяющие П. предыдущих поколений. П., развивающийся из одного очага верхушечной меристемы, у многолетников может расти верхушкой (моноподиально) в течение ограниченного времени или очень долго и притом с перерывами, обусловленными внешними или внутренними причинами (зима, засуха, корреляции в росте отдельных органов). П., вырастающий из почки за один период роста, называется элементарным, а в течение года - годичным. В умеренном климате большинство древесных растений имеет лишь один прирост за год, т. о. элементарный П. у них является и годичным. Но у дуба, например, нередко в середине лета наступает второй период роста и образуются т. н. ивановы П.; тогда годичный П. состоит из двух элементарных. П. цитрусовых дают в год нередко 3-4 прироста, тропические деревья (какао, бразильская гевея) - до 7 приростов. Из ряда годичных П. сложены многолетние скелетные П. древесных растений. Длина междоузлий в пределах элементарного или годичного П. отражает изменение интенсивности его роста: у основания П. междоузлия обычно короткие, затем более длинные, а к верхушке опять короткие (например, у рябины, жимолости, черёмухи, щавеля, тысячелистника). У некоторых растений (например, у липы, вяза, орешника) годичный П. заканчивается наиболее длинным междоузлием в результате недоразвития и раннего отмирания его верхушечной части. Недоразвитие междоузлий и сближение листьев приводят к образованию розеточных П. Длина междоузлий определяет и тип П. - ростовых, или удлинённых, и укороченных. Последние у деревьев часто специализированы как цветоносные ("плодушки"); зелёные листья на них немногочисленны или недоразвиты (например, у вишни, миндаля, вяза, волчьего лыка). У трав, наоборот, цветоносные побеги, как правило, удлинённые. У луговых трав удлинённые П. составляют основную массу сена, а укороченные - основу пастбищного корма; у плодовых деревьев и кустарников удлинённые П. образуют скелет кроны, а укороченные приносят плоды. В зависимости от цели выращивания древесных растений можно регулировать соотношение удлинённых и укороченных П. их обрезкой.
По продольной оси П. закономерно изменяются величина и форма листьев: нижние нередко чешуевидные, срединные - зелёные ассимилирующие, верхние - прицветные. Это различие (Гетерофиллия) определяется изменением возрастного состояния точки роста П. и условий в период формирования листьев. В формировании и росте П. различают две фазы: эмбриональную, или внутрипочечную, в течение которой идёт заложение новых органов П., и постэмбриональную, или внепочечную, когда развёртываются и растут уже заложенные органы и иногда формируются новые. Если к зиме в почке многолетнего растения заложены все элементы будущего годичного П., включая соцветия и цветки, то весной происходит лишь их развёртывание (у большинства деревьев и кустарников умеренного пояса, у раноцветущих многолетних трав). Если же П. заложен в зимующей почке лишь частично, то весной и летом наряду с развёртыванием уже заложенных элементов годичного П. идёт образование новых его метамеров (у водяных побегов деревьев и кустарников, у поздноцветущих трав). У однолетников рост П. идёт преимущественно за счёт элементов, новообразующихся на конусе нарастания.
Различают П. ортотропные (см. Ортотропизм), растущие вертикально вверх или, как исключение, вниз, и П. плагиотропные (см. Плагиотропизм) - горизонтальные или наклонные. У травянистых растений П. часто анизотропны, т. е. меняют направление роста: вначале растут горизонтально, затем изгибаются и растут вертикально. Заложение соцветий при этом начинается, как правило, лишь при переходе П. в ортотропное положение.
Развитие П. многолетних трав, как и однолетних, обычно завершается формированием соцветий и цветков, однако после цветения и плодоношения П. у многолетников не отмирают полностью, сохраняются их базальные участки, несущие почки возобновления. Цикл развития такого монокарпического П. от раскрытия почки до плодоношения может длиться 1 вегетационный период (моноциклический П. - у иван-чая, купены, норичника), 2 года (дициклический П. - у медуницы), 3 и более лет (три-и полициклический П. - у ковылей, щучки, грушанок).
Кроме типичных ассимилирующих надземных П., у растений образуются разнообразные формы метаморфизированных П. (см. Метаморфоз) со специфическими структурно-биологическими особенностями, связанными с выполнением ими функций запасания, возобновления, вегетативного размножения, иногда - защиты (колючки), лазания (усики). К числу метаморфозов П. относят и образование цветка (См. Цветок) как органа семенного размножения. Многообразие типов П., определяющее жизненные формы (См. Жизненная форма), растений, возникло в процессе длительной эволюции как приспособление к разнообразию условий их жизни, а у культурных растений - и под воздействием человека.
Лит.: Серебряков И. Г., Морфология вегетативных органов высших растений, М., 1952; его же, Экологическая морфология растений, М., 1962; Мейер К. И., Морфология высших растений, М., 1958; Синнот Э., Морфогенез растений, пер. с англ., М., 1963; Первухина Н. В., Проблемы морфологии и биологии цветка, Л., 1970; Zimmermann W., Die Telomtheorie, Stuttg., 1965; Lehrbuch der Botanik, 30 Aufl., Jena, 1971.
Г. И. Серебрякова.
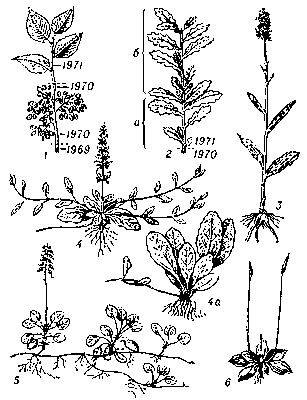
Побеги: 1 - схема побегов вяза. Удлиненный вегетативный побег состоит из 2 годичных побегов, прошлогодний побег уже безлистный (на нём боковые укороченные цветочные побеги); 2 - годичный побег дуба, состоящий из 2 элементарных побегов, 2-й прирост (б) - иванов побег; 3 - моноциклический удлинённый побег у ятрышника; 4 - диполициклические побеги живучки ползучей. Вверху - ортотропный полурозеточный цветоносный побег и плагиотропные боковые столоны. Внизу (4а) - образование ортотропной розетки на конце столона. Розеточный побег зацветает через 1-2 года; 5 - полициклические побеги у грушанки; 6 - розеточный побег подорожника с неполным циклом и безлистные пазушные цветоносы-стрелки.
II
Побе́г
по советскому уголовному праву преступление, выражающееся в уклонении от отбывания наказания (См. Наказание) или меры пресечения (См. Мера пресечения) в виде заключения под стражу. По УК РСФСР побег из места заключения или из-под стражи, совершенный лицом, отбывающим наказание или находящимся в предварительном заключении, наказывается лишением свободы на срок до 3 лет; П., соединённый с насилием над стражей, - до 5 лет; П. с места ссылки, из лечебно-трудового профилактория (либо с пути следования в ссылку или профилакторий) наказывается лишением свободы на срок до 1 года.
ПОБЕГ
СТРАНИЦА ЗНАЧЕНИЙ В ПРОЕКТЕ ВИКИМЕДИА
Побег (фильм); Побег (фильм, 2005)
I
бегство из места заключения, из плена, тайный уход.
П. из тюрьмы.
II
молодая ветка, стебель растения с листьями и почками.
Зеленые побеги.
Wikipedia
Список эпизодов телесериала «Побег»
Список эпизодов американского телесериала «Побег», выходившего на телеканале Fox с 29 августа 2005 года по 15 мая 2009 года. В 2017 году вышло продолжение сериала.

