Введите слово или словосочетание на любом языке 👆
Язык:
Перевод и анализ слов искусственным интеллектом ChatGPT
На этой странице Вы можете получить подробный анализ слова или словосочетания, произведенный с помощью лучшей на сегодняшний день технологии искусственного интеллекта:
- как употребляется слово
- частота употребления
- используется оно чаще в устной или письменной речи
- варианты перевода слова
- примеры употребления (несколько фраз с переводом)
- этимология
Что (кто) такое ДИНОЗАВРЫ: ПТИЦЕТАЗОВЫЕ - определение
ОТРЯД ДИНОЗАВРОВ
Ornithischia; Птицетазовые динозавры; Орнитисхии







.svg?width=200 "30px")













![[[Пентацератопс]]](https://commons.wikimedia.org/wiki/Special:FilePath/Pentaceratops BW.jpg?width=200 "[[Пентацератопс]]")
![[[Игуанодон]]](https://commons.wikimedia.org/wiki/Special:FilePath/Iguanodon new NT.jpg?width=200 "[[Игуанодон]]")
![[[Ламбеозавр]]](https://commons.wikimedia.org/wiki/Special:FilePath/LambeosaurusDB.jpg?width=200 "[[Ламбеозавр]]")
Найдено результатов: 25
ДИНОЗАВРЫ: ПТИЦЕТАЗОВЫЕ
К статье ДИНОЗАВРЫ
Этот монофилетический отряд более разнообразен по составу, чем ящеротазовые. Все птицетазовые были строго растительноядными и в целом эволюционно более продвинутыми, чем рассмотренные выше динозавры. Специализация затронула как фундаментальные особенности их строения, например форму таза, ставшего похожим на птичий, так и адаптации, связанные с питанием, передвижением и защитой от врагов. Наибольшего процветания группа достигла в меловом периоде, что совпало с распространением цветковых растений. Впрочем, хотя птицетазовые и доминировали среди наземных рептилий в позднем мелу, важной частью фауны были в это время также ящеротазовые хищники, страусоподобные динозавры, некоторые завроподы и птицы.
Традиционно выделяли 4 сравнительно слабо различающиеся по уровню организации группы птицетазовых. К орнитоподам ("птиценогим") относятся как примитивные, так и более специализированные двуногие рептилии. Три другие группы образованы высоко специализированными растительноядными формами, передвигавшимися на четырех ногах. Это преимущественно юрские стегозавры (пластинчатые динозавры), теловые анкилозавры (панцирные динозавры) и цератопсы (рогатые динозавры). Кроме похожего на птичий таза, для них характерно присутствие в нижней челюсти беззубой предзубной кости, расположенной спереди от типичной для всех динозавров зубной, несущей зубы. Впрочем, на обеих челюстях передних зубов, как правило, не было, и оканчивались они клювом. Зубы находились только в щечной области и были преобразованы для перетирания и разрезания растительного корма. В отличие от завропод, у некоторых из этих групп зубной аппарат высокоспециализированный и довольно мощный. Общие особенности таза и челюстей свидетельствуют, что все птицетазовые, несмотря на их различия, произошли от общего предка.
В то же время, как показал кладистический анализ, традиционное разделение этого отряда на 4 группы неудовлетворительно. Стегозавров и анкилозавров теперь объединяют в таксон Thyreophora. В его рамках к наиболее архаичным, возможно, наряду со скорее всего двуногим Scutellosaurus, относятся сцелидозавриды, снабженные легким панцирем формы, передвигавшиеся на четырех ногах. Они появились приблизительно на рубеже триаса и юры. Однако еще примитивнее триасовые роды Lesothosaurus (Южная Африка), Technosaurus (Северная Америка) и Pisanosaurus (Южная Америка). Их родственные связи с остальными птицетазовыми пока не вполне выяснены. Орнитоподы, ранее считавшиеся наиболее примитивными из птицетазовых, по-видимому, эволюционно более продвинуты, чем древнейшие тиреофоры. Последними появились цератопсы, включавшие протоцератопсов и собственно цератопсов, иногда называемых неоцератопсами. "Попугаеклювые" пситтакозавры и купологоловые пахицефалозавры связаны общими предками с цератопсами.
Тиреофоры. Эта группа включает несколько типов довольно примитивных панцирных форм. Их объединяет несколько общих признаков черепа, а также присутствие на теле продольных рядов килеватых костных щитков. Как уже говорилось, к архаичным родам этой группы относится Scelidosaurus, однако гораздо лучше изучены стегозавры и анкилозавры.
Стегозавры появились в начале юрского периода и представляют собой самых ранних хорошо известных птицетазовых. Сам род Stegosaurus относится к верхней юре. Это был динозавр длиной более 6 м, с двумя рядами очень крупных костных пластин, вертикально в шахматном порядке поднимавшихся по бокам от позвоночника. Хвост ближе к концу был усажен мощными шипами, а в области шеи на коже располагались костные бляшки. Назначение спинных пластин не вполне ясно. Предположение о возможной теплокровности динозавров привело к пересмотру распространенного прежде мнения, согласно которому речь идет о защитных образованиях. Эксперименты с моделями в аэродинамических трубах показали, что эти пластины могли служить для охлаждения тела, действуя в качестве отражательных экранов, направлявших потоки воздуха вокруг своих поверхностей. Такую точку зрения подтверждают данные о наличии в них густой сети каналов для кровеносных сосудов.
Стегозавры передвигались на четырех ногах, хотя их передние конечности гораздо короче и легче задних. Голова крайне мала по сравнению с общими размерами тела. Спинной мозг у них, как и у некоторых других динозавров, утолщался в тазовой области, что привело к популярному, хотя и ошибочному представлению, будто у стегозавров было как бы два "головных" мозга. На самом деле "второй мозг" управлял лишь двигательной активностью массивных задних конечностей и хвоста. Стегозавры жили в тех же сообществах, что и крупные позднеюрские завроподы.
Анкилозавры (панцирные динозавры) составляют крайне интересную, но недостаточно изученную группу птицетазовых. Известны они прежде всего из меловых отложений Северной Америки, Европы, Азии и, возможно, Южной Америки. Их остатки немногочисленны, и эволюционная история остается неясной. Эти динозавры удачно названы "рептилиями-танками", их крупные представители достигали в длину 6 м. У типичных анкилозавров из родов Nodosaurus, Ankylosaurus и Palacoscincus панцирь устроен следующим образом. Чередующиеся ряды крупных и мелких многоугольных пластин формировали покрывающую все тело защитную мозаику, на ногах были шипы, на хвосте - панцирные кольца, в некоторых случаях с шипами, а на голове как бы сплошной костный шлем. Форма туловища тяготела к широкой и уплощенной, а держалось оно на коротких мощных конечностях. Находящиеся в Москве находки из Монголии позволяют проследить ряд форм, сменявших друг друга с нижнего мела до конца этого периода. Лучше всего изучен Syrmosaurus, внешним видом несколько напоминающий многократно увеличенную жабовидную ящерицу. Самым крупным был Ankylosaurus, достигавший в длину 10,7 м.
Большинство анкилозавров было медлительными фитофагами. Их слабые челюсти и почти отсутствующие у некоторых форм зубы указывают на питание мягкими частями растений. Об их происхождении почти ничего не известно, хотя некоторые формы из раннего мела Европы несколько примитивнее типичных и лучше изученных родов.
Орнитоподы. Среди древнейших остатков птицетазовых изредка встречаются "птиценогие" ящеры, датируемые поздним триасом или самым началом юры. Однако немногие хорошо сохранившиеся образцы и некоторые фрагментарные остатки дают достаточно ясное представление об этих животных. Лучше других известен Heterodontosaurus, двуногое существо длиной ок. 0,9 м, а также род Fabrosaurus. Судя по ним, в конце триаса уже существовало по крайней мере два четко обособленных семейства орнитопод, следовательно, вся эта группа оформилась не позднее среднего триаса. Некоторые архаичные ее представители, т.н. эволюционные реликты, встречаются и в отложениях мелового возраста. Таков, например, род Hypsilophodon из нижнего мела. Это было маленькое быстро бегающее двуногое животное, примитивность которого выражалась в сохранении передних зубов на верхней челюсти. Хвост его длинный и тонкий, а вытянутые пальцы задних ног, вероятно, хорошо гнулись. Сравнительно примитивная юрская форма Camptosaurus дает представление о промежуточной эволюционной стадии между ранними и более специализированными орнитоподами. Это был тяжелый ящер с довольно крупными передними конечностями, позволявшими предположить, что передвигался он в основном на 4 ногах. Зубы и челюсти хорошо приспособлены для питания растительным кормом, а беззубый клюв служил, вероятно, для скусывания сочных побегов.
Более специализированны "утконосые" орнитоподы, называемые траходонтами или гадрозаврами. У них был плоский клюв без передних зубов. Однако в щечной области челюсти несли мощные перетирающие зубы, хорошо приспособленные к питанию грубой растительной пищей. Утконосые динозавры были крупными, тяжеловесными животными. Длина их тела в среднем превосходила 9 м. Задние конечности были массивными, а передние хотя и более короткими, но хорошо развитыми. Обычно они передвигались на двух ногах, но могли становиться и на четвереньки.
Среди утконосых динозавров обнаружены колоссальные различия в строении черепа, касающиеся в первую очередь окружающих ноздри костей. У некоторых они образуют причудливые вздутия в носовой области, у других полые куполообразные гребни или длинные выступающие позади головы трубчатые структуры.
По-видимому, в конце мелового периода утконосые динозавры были распространены почти по всему свету. В Северной Америке они лучше всего известны из Канады, где представлены как консервативные формы, например Anatosaurus, так и специализированные типы вроде Lambeosaurus с высоким вздутым гребнем на голове. Их остатки встречаются главным образом в осадочных породах, сформировавшихся на дне рек, озер и болот. Вероятно, эти животные вели амфибиотический образ жизни и хорошо плавали. Известны также их мумифицированные остатки с сохранившимся рельефом кожи и очертаниями формы тела. Они показывают, что тело покрывали мелкие чешуи, панциря не было.
Цератопсы, или рогатые динозавры, появились среди гигантских рептилий последними. Их череп оканчивался высоким и узким беззубым клювом, а сзади разрастался в широкий костный воротник, который нависал над шеей примерно настолько же, насколько простиралась вперед остальная часть головы. Известно несколько вариантов "рогатости" этих рептилий. Некоторые из них практически безроги, другие несли лишь один рог - на рыле, а, скажем, у рода Triceratops их было три - по одному на рыле и над глазами. Традиционно считается, что эти рога использовались в схватках с хищниками. Однако аналогия с ныне живущими рогатыми животными, например буйволом, наводит на мысль об определенной роли таких структур в распознавании полового партнера и других формах социального поведения, особенно связанных с доминированием и спариванием.
Задние ноги цератопсов длинные и тяжелые, а передние, хотя и не столь внушительные, для птицетазовых относительно крупные; лапы широкие и короткие. Туловище было толстым, а хвост массивным и коротким, приблизительно равным ему по длине. Панцирь, если не считать головного воротника, отсутствовал.
Цератопсы известны только из мела, однако они успели дать множество разнообразных морфологических типов. Protoceratops из Монголии - наиболее известная мелкая примитивная форма. На серийном материале, собранном экспедицией Американского музея естественной истории, изучен процесс индивидуального роста этих ящеров, в частности развития воротника или гребня на заднем крае их черепа. Сначала его разрастание увеличивало площадь крепления мышц, приводящих в движение нижнюю челюсть. На более поздних стадиях, а также по мере специализации цератопсов воротник, вероятно, принимал на себя функцию защиты шеи как наиболее уязвимой области тела. С учетом данных, указывающих на возможную теплокровность этих ящеров, т.е. их способность поддерживать постоянную температуру тела, можно предположить, что воротник участвовал и в ее контроле, при избытке тепла рассеивая его, а при недостатке - улавливая из внешней среды.
Особенно много разнообразных цератопсов обнаружено в верхнемеловых отложениях Азии и Северной Америки. Больше других известен Triceratops длиной ок. 6 м. Его череп нес огромный воротник и три хорошо развитых рога. Ноздри были очень велики, верхняя челюсть завершалась мощным клювом, щечные зубы развивались слабо. Несколько меньший Monoclonius отличался присутствием всего одного рога на морде. У рода Styracosaurus тоже был лишь один носовой рог, зато его воротник окаймляли длинные шипы.
Цератопсы относятся к наиболее многочисленным динозаврам конца мелового периода. Вероятно, они были хорошо адаптированы к жизни на открытых пространствах и питанию цветковыми растениями. Несмотря на обилие этих ящеров, прежде всего из рода Triceratops, они не пережили перехода от мезозоя к кайнозою, и их вымирание ознаменовало конец эпохи динозавров.
В начале 1920-х годов в нижнемеловых отложениях монгольской пустыни Гоби обнаружены гнезда с яйцами Protoceratops. Со времени этого открытия было найдено много других гнезд динозавров. Прекрасно сохранившиеся захоронения такого типа раскапывают в Монголии. Они содержат целые кладки яиц утконосых динозавров с невылупившимися эмбрионами и скелеты их детенышей. Там же найдены гнезда мелких двуногих орнитопод и неизвестных крупных животных.
Как показали эти находки, уход за гнездом и забота о потомстве у утконосых динозавров были сложнее, чем у современных крокодилов, которые обнаруживают лишь зачатки родительского поведения. Такие особенности, как строительство гнезда, признаки заботы о потомстве вместе с возможной теплокровностью, широким разнообразием типов динозавров и освоенных ими местообитаний, а также свидетельствами их стадного поведения, позволяют говорить о сложной социальной структуре сообществ этих мезозойских рептилий, развивавшейся на протяжении 100 млн. лет их господства. Возможно, по уровню поведенческой организации они приближались к современным птицам или млекопитающим.
Птицетазовые динозавры
(Огnithischia)
отряд ископаемых пресмыкающихся надотряда динозавров (См. Динозавры). Имели четырёхлучевое строение таза с развитым задним отростком лобковой кости, как у птиц. Передняя, лишённая зубов часть челюстей была одета роговым чехлом и имела вид клюва. Зубы сжатые с боков, с листовидными коронками. Концевые фаланги пальцев у поздних П. д. уплощены в виде копыт. Были растительноядны (самые ранние, возможно, - всеядны). Многие обладали для защиты от хищников различными приспособлениями в виде кожных щитков, шипов или рогов. 6 (4) подотрядов, в том числе Стегозавры, Анкилозавры и Рогатые динозавры. Жили в юрский и меловой периоды.
динозавр
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
ДИНОЗАВР
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
вымершее крупное пресмыкающееся.
Динозавры
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
(Dinosauria)
самый многочисленный надотряд ископаемых мезозойских пресмыкающихся подкласса архозавров (См. Архозавры). Делят на 2 отряда: ящеротазовые и птицетазовые Д., которые включают 7 подотрядов, объединяющих несколько сот видов. Самые ранние Д. известны со среднего триаса, последние Д. дожили до конца мелового периода. Д. - морфологически и экологически крайне разнообразная группа. Предками Д. были более примитивные триасовые архозавры - Текодонты, от которых Д. унаследовали способность к передвижению на двух задних ногах (бипедализм), со временем у одних усилившуюся (хищные Д. и Орнитоподы), а у других - утраченную (зауроподы, стегозавры, анкилозавры и рогатые Д.). Первично все Д. были сухопутными хищниками, но уже в конце триаса от них отделились Прозауроподы, возможно, всеядные Д., давшие начало гигантским растительноядным зауроподам (См. Зауроподы), перешедшим к обитанию в крупных внутриматериковых водных бассейнах или в прибрежной зоне моря. В триасе же появились гетеродонтозавры (с передними зубами, как у хищных Д., а задними, как у орнитопод), давшие, вероятно, начало всем птицетазовым Д. - растительноядным животным. У большинства из них (исключая орнитопод, ставших, как и зауроподы, обитателями водоёмов) развились защитные приспособления против хищных Д. (шипы и костные панцири у стегозавров (См. Стегозавры) и анкилозавров, рога - у рогатых Д.). Размножались Д. яйцами, скорлупу которых находят довольно часто. Возможно, Д. были теплокровными животными, обладали и др. признаками высокой физиологической организации. Остатки Д. - многочисленные скелеты, черепа и др. встречаются на всех материках; в СССР - главным образом на территории Казахстана, а также Таджикистана и Киргизии (район Ферганы). Изучение Д. важно для познания эволюции позвоночных. Широкое географическое распространение и быстрая смена видов Д. во времени делают их ценными руководящими формами для определения геологического возраста континентальных отложений мезозоя. На протяжении всего мезозоя на смену одним видам, родам, семействам и подотрядам появлялись новые группы. Время исчезновения последних Д. соответствует границе между мезозоем и кайнозоем. Основными причинами вымирания Д. следует считать неблагоприятные изменения среды обитания (перераспределение воды и суши на земной поверхности с возможным исчезновением естественных зон обитания, смена климата, нарушение баланса в составе флоры и фауны, т. е. пищевой базы, и т.д.), вызываемые первично геологическими преобразованиями земной коры, которые были весьма значительны в мезозое, особенно в позднем мелу.
Лит.: Основы палеонтологии, [т. 12]. Земноводные, пресмыкающиеся и птицы, М., 1964; Рождественский А. К., На поиски динозавров в Гоби, 3 изд., М., 1969; Colbert Е. Н., Men and dinosaurs, N. Y., 1968.
А. К. Рождественский.
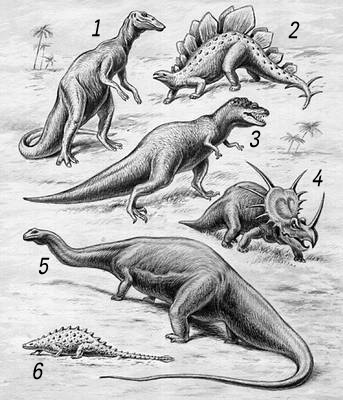
Динозавры: 1 - анатозавр; 2 - стегозавр: 3 - тарбозавр; 4 - стиракозавр: 5 - апатозавр; 6 - талорурус.
ДИНОЗАВРЫ
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
популярное название группы вымерших наземных рептилий, которая включает самых крупных после китов животных, известных науке. Динозавры жили в мезозойскую эру, начавшуюся приблизительно 225 млн. лет назад и закончившуюся ок. 65 млн. лет назад.
Термин "динозавры" (Dinosauria; от греч. deinos - страшный и saurus - ящер) ввел в научную литературу британский палеонтолог Р.Оуэн в 1842. Он назвал так отряд, в который объединил разнообразных рептилий, известных в то время по ископаемым костям из Европы и Северной Америки. С тех пор динозаврами стали называть любых крупных вымерших ящеров. Однако такое расширительное использование термина неправильно, поскольку не все гигантские наземные рептилии мезозойской эры были динозаврами, а некоторые динозавры были по размерам мельче петуха.
В мезозое рептилии доминировали среди позвоночных на суше, играли важную роль в водных экосистемах и даже освоили воздушное пространство. Большинство этих ящеров относят к одной обширной таксономической группе, подклассу Archosauria класса Reptilia. Примитивные архозавры объединены в группу текодонтов. К этому же подклассу относятся крокодилы, летающие ящеры (птерозавры), динозавры, а в некоторых классификационных схемах даже птицы. До наших дней дожили только птицы и крокодилы.
Динозавров принято делить на два отряда - ящеротазовых (Saurischia) и птицетазовых (Ornithischia). Раньше считалось, что они произошли от текодонтных предков независимо друг от друга. Таким образом, слово "динозавр" утрачивало свой первоначальный таксономический смысл и оставалось лишь удобным популярным обозначением вымерших ящеротазовых и птицетазовых рептилий.
Однако более детальный анализ позволил выявить характерный для обоих отрядов уникальный набор признаков, и сейчас принято считать, что все динозавры происходят от одного общего предкового вида, т.е. представляют собой монофилетическую группу, или кладон (ветвь на родословном древе животных).
Ящеротазовые названы так по причине сходства их таза с тазом большинства рептилий. Среди них были как хищные, так и растительноядные формы. В этом отряде, вероятно, лучше всего известны гигантские растительноядные завроподы длиной до 24 м и более. Столь же внушительными были чудовищные двуногие хищники с огромными черепами, челюстями и зубами.
Птицетазовые отличаются формой лобковых костей таза, которые, как и у птиц, несут длинный отросток, направленный более или менее параллельно седалищной кости. У большинства же рептилий лобковая и седалищная кости расходятся под значительным углом. Все птицетазовые динозавры растительноядны.
Многие ранние представители как ящеротазовых, так и птицетазовых были двуногими. Однако некоторые их потомки в обеих группах вновь освоили передвижение на четырех ногах. Эти рептилии господствовали на суше в мезозое ок. 100 млн. лет. К началу нынешней, кайнозойской эры они вымерли, и век рептилий сменился веком млекопитающих.
Находки динозавров. Остатки динозавров найдены во многих районах Северной Америки. На востоке США в долине реки Коннектикут породы триасового возраста (начало мезозойской эры) содержат множество отпечатков ног динозавров. Изучая такие следы, можно многое узнать о том, как они передвигались, какие их формы обитали совместно. Близ Вернала (шт. Юта) в бассейне Уинта раскопано обширное кладбище динозавров юрского периода (середина мезозойской эры). Среди представленных там видов много гигантских растительноядных завропод, самых крупных сухопутных рептилий. Много остатков динозавров близкого возраста найдено в Комо-Блафс (шт. Вайоминг) близ Медисин-Боу. Очень хорошо представлены они в слоях более позднего возраста, относящихся к меловому периоду, т.е. к концу мезозойской эры, в штате Монтана и на западе Канады.
Разумеется, гигантские ящеры - это далеко не все динозавры. Мелкие формы интересны сами по себе, и, что важнее, некоторые из них указывают на связь этой группы с более ранними, архаичнее организованными рептилиями. Примитивные динозавры, самые ранние из которых известны с верхнего триаса (ок. 200 млн. лет назад), лишь продолжили историю пресмыкающихся, которая началась по крайней мере за 150 млн. лет до их времени. Доминирующее положение динозавры стали приобретать в конце триаса. На протяжении двух последующих периодов, юрского и мелового, эта группа процветала, а затем быстро вымерла.
См. также:
Ящеротазовые динозавры
СОГЛАСНО НАИБОЛЕЕ РАСПРОСТРАНËННОЙ КЛАССИФИКАЦИИ, ОДНА ИЗ ДВУХ ОСНОВНЫХ КЛАД ДИНОЗАВРОВ
Saurischia; Ящеротазовые динозавры; Заврисхии; Eusaurischia
(Saurischia)
отряд динозавров (См. Динозавры). Для Я. д. характерно трехлучевое строение тазового пояса (свойственное большинству пресмыкающихся), что отличает их от птицетазовых динозавров (См. Птицетазовые динозавры). Жили от середины триаса до конца мелового периода. По строению и образу жизни делятся на 2 подотряда: хищные двуногие тероподы и растительноядные зауроподы. Я. д. были широко распространены по всему земному шару.
Анкилозавры








![Модель питающегося анкилозавра рода ''[[Euoplocephalus]]''](https://commons.wikimedia.org/wiki/Special:FilePath/Euoplocephalus eating Animatronics model NHM2.jpg?width=200 "Модель питающегося анкилозавра рода ''[[Euoplocephalus]]''")









.jpg?width=200 "Генри Фэрфилд Осборн")
![[[Сцелидозавр]] является ближайшим предком анкилозавров](https://commons.wikimedia.org/wiki/Special:FilePath/Scelidosaurus harrisonii.png?width=200 "[[Сцелидозавр]] является ближайшим предком анкилозавров")




ИНФРАОТРЯД ПТИЦЕТАЗОВЫХ ДИНОЗАВРОВ
Панцирные динозавры; Ankylosauria
(Ankylosáuria)
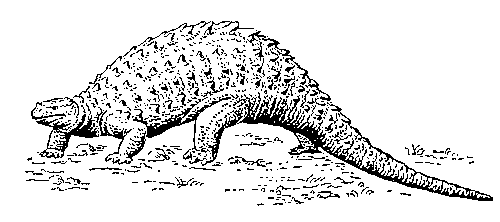
панцирные динозавры, подотряд птицетазовых динозавров, характерных для мелового периода. Крупные (длина до 8-9 м) растительноядные четвероногие пресмыкающиеся. Тело широкое, уплощённое; защищено сверху костными шипами или щитками, слившимися у некоторых А. в сплошной панцирь (отсюда другое название). На конце хвоста, служившего активным средством защиты от хищных динозавров, нередко имелись острые шипы. Остатки А. известны на всех материках (кроме Австралии и Антарктиды), но в основном - из верхнего мела Сев. Америки, Европы и Азии; в СССР - Центральный Казахстан, Кызылкум.
Лит.: Основы палеонтологии, т. 12 - Земноводные, пресмыкающиеся и птицы, М., 1964.
А. К. Рождественский.
Анкилозавр.
Динозавры
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
Динозавры (чудовищные ящеры) - вымерший порядок пресмыкающихся,установленный англ. учен. Овеном, представители которого соединяли всебе свойства нескольких, ныне обособленных групп животного царства:ящериц, крокодилов, млекопитающих и птиц. Череп Д. вообще устроен потипу ящериц, но каждый зуб помещается в отдельной ячейке, как укрокодилов; Подобно последним, туловище у некоторых родов покрытокожистым или костяным панцирем. С млекопитающими Д. сближает строениетрубчатых костей с широкими каналами, выполненными мозговым веществом,строение неуклюжих суставов пальцев и крестцовой кости; наконец построению таза и задних конечностей они более всего приближаются кптицам. Крупные и крепкие кости конечностей, приспособленные для ходьбы,заставляют предполагать, что Д. обитали на суше, в лесистых, болотистыхнизинах, а массивный крестец, крепкий таз и чрезвычайно развитые задниеконечности их более приспособлены к прямой постановке туловища приотдыхе и передвижении, как у современных кенгуру. Д. появляются втриасовый период, пользуются значительным распространением в течениеюрского и безвозвратно угасают в меловом периоде. Наиболее многочисленныостатки их в юрских отложениях Скалистых тор (Сев. Америка), а также впресноводных осадках юрской и меловой системы Англии, Бельгии иГермании. К Д. относятся животные очень различные по наружному виду,образу жизни и анатомическому строению, принадлежащие к 20 родам,включающим несколько десятков видов. Некоторые представители этой группыизвестны только по уцелевшим отпечаткам следов, в то время как от другихнайдены полные, прекрасно сохранившиеся скелеты. Отпечатки следов Д.,сделанные ими на рыхлом песке, отвердевшем впоследствии в песчаник, ужедавно были найдены на плитах триасового песчаника Коннектикута (Сев.Америка) под именем орнитихнитов и приписывались долгое времяисполинским птицам. Теперь, однако, доказано, что эти многочисленныетрехпалые парные следы принадлежат по крайней мере 30 различным видамД., от которых в этих отложениях не уцелело ни одной кости скелета. Повеличине Д. очень различны: одни, как, напр., компсогнат, были величинойс кошку; другие являлись гигантами, превосходящими своими размерами всехдругих, когда либо попиравших земную поверхность животных; таков, напр.,атлантозавр. По характеру зубного вооружения можно судить об образежизни и пище Д. Одни из них (мегалозавр, компсогнат) принадлежали кплотоядным хищникам, другие (как игуанодон) - к травоядным. К хищным Д. относится, между прочим, один из наиболее древнихпредставителей этой группы, цанклодон, от которого сохранился почтиполный скелет из триасовых отложений Виртемберга. Этот гигантскийхищник, снабженный сильными, почти одинаково развитыми пятипалымиконечностями, достигал 9 - 10 м в длину и был вооружен многочисленныминожевидными зазубренными зубами. О величине цанклодона можно судитьпотому, что отпечатки ступни этого животного занимают площадь в 3 кв.фт. В юрских отложениях Англии и Германии довольно часто встречаютсяостатки другого хищника - мегалозавра, очень немного уступавшегопредыдущему по величине. Но наибольший интерес в этой группепредставляет грациозный компсогнат, полный скелет которого найден взнаменитом, по многочисленным интересным находкам окаменелостей,золенгофенском сланце Баварии. Компсогнат был небольшой зверёк,величиной с домашнюю кошку. При взгляде на его скелет прежде всегобросается в глаза неравномерное развитие передних и задних конечностей,из которых только последние служили для передвижения скачками, придаваяэтому животному сходство с современными тушканчиками и кенгуру.Приближаясь по строению черепа к ящерицам, компсогнаты, по устройствутаза и задних конечностей, очень сходны с птицами. На передних и заднихногах у них было по три пальца. Длинный и сильный хвост, на который ониопирались, значительно помогал им при передвижении. В числе травоядных Д. наиболее древним считается крупный наземныйящер сцелидозавр, также с неравномерно развитыми конечностями, одетыйпанцирем, состоящим из многих костяных пластинок. К травоядным относитсяи наибольший не только из Д., но и изо всех наземных животныхатлантозавр, описанный Маршем из юрских отложений Скалистых гор С.Америки, диаметр бедренной кости которого равняется 63 стм., а общаядлина тела доходит до 40 м. Передние и задние конечности его развитыпочти одинаково. Этот исполин населял болотистые низменности, питаясь,по всем вероятиям, листьями деревьев. В тех же Скалистых горах найденыполные скелеты еще двух травоядных Д.: близкого к предыдущему поанатомическому строению, но значительно меньших размеров (10 - 12 м.)морозавра, и чудовищного бронтозавра, достигавшего 16 м в длину.Последний, как предполагают, водный обитатель, был снабжен чрезвычайнодлинным хвостом и такой же шеей, на которой сидела ничтожная по размерамголовка. Крепкие конечности, из которых задние развиты несколько болеепередних, снабжены пятью пальцами, вооруженными когтями. К числутравоядных Д. принадлежит также ранее других открытый и наилучшеизученный неуклюжий игуанодон. Кости последнего еще в 1818 г. былизамечены Мантеллем в пресноводных вельдских отложениях Англии, изкоторых впоследствии были собраны многочисленные остатки его скелетов инайдены отпечатки трехпалых задних конечностей; но только в 1878 г.,когда в Бельгии, в Берниссарте, было добыто до 20 полных скелетовигуанодона, получили о нем полное представление. Это гигантское животноедостигало 10 метров в длину и до 4,5 м. в вышину, в свойственном емуполустоячем положении. Подобно большинству Д. и игуанодон передвигалсятолько на двух задних массивных конечностях, подпираясь длинным исильным хвостом. Передние конечности, развитые гораздо менее, снабжены 5пальцами, тогда как на задних вполне развито только 3 пальца, ачетвертый является в форме шпоры. Таз, несмотря на громадные размеры,приближается по строению к тазу птиц. Многочисленные зубы игуанодонаимеют вид терки, которой животное перетирало растительную пищу, а каждыйзуб в отдельности представляет складчатую, по краям зазубреннуюлопаточку, очень напоминая зубы ныне живущей ящерицы игуаны, отчего этиживотные и получили свое название. По строению конечностей и наружныхпокровов в настоящее время, следуя Маршу, Д. разделяют на несколькогрупп, из которых главнейшие: Sauropoda(ящеронoгие), Stegosauria(панцирные Д.), Ornitopoda (птиценогие) и Theropoda (хищные). Основныеработы о Д.: Marsch. "Principal characters of American jurassicDinosauria" и др. статьи в "Americ. Journ" (Т. XVI - XXXIX); Rich. Owen,"A monograph on the Reptilia of the Wealden and Purbeck formations"("Paleont soc.", t. I - V, and Suppl. I - IX) и др. ст. в том же изд.;Seeley. "On the classification of the Dinosauria" ("Proceed. Roy. Soc.",1887). Б. П.
ДИНОЗАВРЫ
КЛАДА АРХОЗАВРОВ
Динозавр; Dinosauria; Нептичьи динозавры; Ужасный ящер; Avipluma
самый многочисленный надотряд вымерших пресмыкающихся. Жили в мезозое, вымерли в конце мелового периода. Длина от 20 см до 30 м. Одни (хищные динозавры, орнитоподы) передвигались на двух ногах, другие (стегозавры, рогатые динозавры) - на четырех. Ок. 600 видов, относящихся к 2 отрядам: ящеротазовые динозавры (апатозавры, диплодоки) и птицетазовые динозавры (игуанодонты, утконосые динозавры, стегозавры, зауролофы). Древние динозавры были сухопутными хищниками. Затем появились растительноядные, некоторые из них перешли к жизни в воде. Руководящие ископаемые.
Википедия
Птицетазовые

Птицета́зовые, или птицета́зовые диноза́вры, или орнити́схии (лат. Ornithischia), — одна из двух основных клад динозавров, рассматриваемая в ранге отряда. Несмотря на своё название, не они, а ящеротазовые динозавры по самой распространённой версии дали начало птицам, у которых «птичий» таз сформировался совершенно независимо.

