Введите слово или словосочетание на любом языке 👆
Язык:
Перевод и анализ слов искусственным интеллектом ChatGPT
На этой странице Вы можете получить подробный анализ слова или словосочетания, произведенный с помощью лучшей на сегодняшний день технологии искусственного интеллекта:
- как употребляется слово
- частота употребления
- используется оно чаще в устной или письменной речи
- варианты перевода слова
- примеры употребления (несколько фраз с переводом)
- этимология
Что (кто) такое ПАЛЕОНТОЛОГИЯ: ТРИАС - определение
ПЕРВЫЙ ГЕОЛОГИЧЕСКИЙ ПЕРИОД МЕЗОЗОЙСКОЙ ЭРЫ
Триас; Триасовая система; Вёрфенский ярус; Триас (геология)

![[[Мастодонзавр]] — гигантский среднетриасовый лабиринтодонт](https://commons.wikimedia.org/wiki/Special:FilePath/Mastodonsaurus DB.jpg?width=200 "[[Мастодонзавр]] — гигантский среднетриасовый лабиринтодонт")
")
![лабиринтодонт]]](https://commons.wikimedia.org/wiki/Special:FilePath/Sclerothorax DB2.jpg?width=200 "лабиринтодонт]]")
Найдено результатов: 21
ПАЛЕОНТОЛОГИЯ: ТРИАС
К статье ПАЛЕОНТОЛОГИЯ
35
225
Саговники, возникновение цветковых растений. Первые млекопитающие, пре-смыкающиеся, включая динозавров, костные рыбы. Саговники и хвойные.
ТРИАСОВАЯ СИСТЕМА
(ПЕРИОД) (триас) (от греч. trias - троица), 1-я (нижняя) система мезозойской эратемы, соответствующая самому раннему периоду мезозойской эры геологической истории. Начался триасовый период 235 ± 10 млн. лет назад, продолжительность 50 млн. лет. Подразделяется на 3 отдела. Для триасового периода характерно обновление морской и наземной фауны. В морях главную роль среди беспозвоночных играли аммоноидеи (цератиты), пелециподы, гастроподы; впервые появились белемниты, костистые рыбы. Характерен расцвет пресмыкающихся - крупных рептилий (динозавров), появились первые млекопитающие (яйцекладущие и сумчатые). В растительном мире преобладали папоротники, цикадофиты, гинкговые и хвойные. Основные полезные ископаемые триасового периода - угли, нефть, алмазы, уран, медно-никелевые руды.
Триасовая система
(период)
--------------------------------------------------------------------------------------------------------------------------------------------------------
| Альпы | Средняя Европа | |
| ------------------------------------------------------------------------------------------------ ------------------------------------------------ ------ |
| Верхний | Рэтский | Верхний кейпер | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | ----------------------------------------------- | | ------ |
| | Норийский | ----------------------------------------------- ------- |
| | | Средний кейпер | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | ----------------------------------------------- | | ------ |
| | Карнийский | | ------ |
| | | | ------ |
| | | | ------ |
| | | ----------------------------------------------- ------- |
| | | Нижний кейпер | ------ |
| ----------------------------------------------- ------------------------------------------------ | | ------ |
| Средний | Ладинский | | ------ |
| | | | ------ |
| | | ----------------------------------------------- ------- |
| | | Раковинный известняк | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | ----------------------------------------------- | | ------ |
| | Анизийский | | ------ |
| | | ----------------------------------------------- ------- |
| | | Верхний пёстрый песчаник | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| ----------------------------------------------- ------------------------------------------------ | | ------ |
| Нижний | Скифский | ----------------------------------------------- ------- |
| | | Средний пёстрый песчаник | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | | ----------------------------------------------- ------- |
| | | Нижний пёстрый песчаник | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
| | | | ------ |
--------------------------------------------------------------------------------------------------------------------------------------------------------
--------------------------------------------------------------------------------------------------------------------------------------------------------------
| Ярусы | Подъярусы | Западная Европа | Северная Америка | Азия | |
| | | -------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | | Альпы (Л. Кристин, В. | Канада и запад США | северо-восток СССР | |
| | | Шёльнбергер, 1972) | (Э.Т. Тсзер, 1971) | (А.С. Дагис и | |
| | | | | др.,1974) | |
| -------------------- -------------------- --------------------------------------- ------------------------------------ ------------------------------------ ----- |
| Рэт | Рэт | Choristoceras marshi | Choristoceras marshi | Tosapecten efimovae | |
| -------------------- -------------------- --------------------------------------- ------------------------------------ | | ---- |
| Норийский | Севат | Rhabdoceras suessi | upper | ----------------------------------- ----- |
| | | | Rhabdoceras suessi | Monotis ochotika | |
| | | | lower | | |
| | | | | | |
| | ------------------- --------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | Алаун | Горизонт с Halcrites | Himavatites | Monotis scutiformis | |
| | | | columbianus | | |
| | | -------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | | Cyrtopleurites bicrenatus | Drepanites rutherfcrdi | Otapiria ussuriensis | |
| | ------------------- --------------------------------------- ------------------------------------- | | ---- |
| | Лак | Juvavites magnus | Juvavites magnus | | |
| | | -------------------------------------- ------------------------------------- | | ---- |
| | | Malayites paulckei | Malayites dawsoni | | |
| | | -------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | | Mojsisovicsites kerri | Mojsisovicsites kerri | Pinacoceras | |
| | | | | verchojannicus | |
| -------------------- -------------------- --------------------------------------- ------------------------------------ ------------------------------------ ----- |
| Карнийский | Туваль | Anatropires - Bereich | Klamathites | Sirenites yacutensis | |
| | | | macrolobatus | | |
| | | -------------------------------------- ------------------------------------- | | ---- |
| | | Tropites subbulatus | Tropites welleri | | |
| | | -------------------------------------- ------------------------------------- | | ---- |
| | | ? | Tropites dilleri | | |
| | ------------------- --------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | Юль | -------------------------------------- | Sirenites nanseni | Sirenites hayesi | ---- |
| | | Trachyceras aonoides | | | |
| | ------------------- --------------------------------------- ------------------------------------- ------------------------------------ ----- |
| | Кордеволь | | | Protrachyceras | |
| | | | | seimkanense | |
| | | Trachyceras aon | Trachyceras obesum | ----------------------------------- ----- |
| | | | | Protrachyceras | |
| | | | | omkutchanicum | |
--------------------------------------------------------------------------------------------------------------------------------------------------------------




триас, первая (нижняя) система мезозойской группы, соответствующая первому периоду мезозойской эры истории Земли; она следует за пермской системой и предшествует юрской. Начало Т. п., по радиологичческим данным, 230 млн. лет назад, конец - 195 млн. лет назад; продолжительность составляет около 35 млн. лет. Название происходит от греческого слова trias - троица, так как Т. с. возникла в результате объединения трёх выделенных в разрезах Центральной Европы комплексов слоев: пёстрого песчаника, раковинного известняка и пёстрого мергеля (Кейпера). Бельгийский учёный Омалиус д'Аллуа в 1831 объединил их под названием "кейперские отложения", а немецкий учёный Ф. Альберти в 1834 - под названием "триас".
На территории СССР отложения Т. с. впервые были установлены Л. Бухом на основании определений в 1831 триасовых цератитов в Прикаспии (из коллекции русского геолога П. С. Палласа, 18 в.); затем триасовая фауна С.-В. была описана русским учёным А. Кайзерлингом (середина 19 в.). Континентальные отложения Т. с. были изучены в конце 19 - начале 20 вв. русскими геологами С. Н. Никитиным, Н. Н. Яковлевым, Н. Г. Кассиным, А. Н. Мазаровичем и др. Большой вклад в расчленение морского триаса внесли советские геологи (В. Н. Робинсон - Кавказ; Е. М. Люткевич, Е. И. Соколова и др. - Восточно-Европейская платформа; Л. Д. Кипарисова, Ю. Н. Попов, И. И. Тучков С.-В. и Дальний Восток; В. И. Славин - Карпаты; В. И. Дронов - Памир). Из зарубежных геологов стратиграфию Т. с. разрабатывали Э. Зюсс, Э. Мойсисович, В. Вааген, К. Динер, А. Битнер, Д. Штур и другие.
Подразделения. Впервые отложения Т. с. были изучены ещё в середине 18 в. на территории Средней Европы - Германии и С.-В. Франции (так называемый Германский бассейн), но выделенные там подразделения из-за их своеобразия и бедности органическими остатками плохо прослеживаются за её пределами. Значение эталона для среднего и верхнего триаса со 2-й половины 19 в. стал играть разрез морских отложений Альп, а для нижнего - разрезы Гималаев и Соляного Кряжа. Т. с. сохранила первоначальное деление на три отдела, хотя отделы альпийского триаса (табл. 1) не являются точными эквивалентами пёстрого песчаника, раковинного известняка и кейпера Германского бассейна. Геологическое строение Северных Известняковых Альп, где находятся стратотипы анизийского яруса (См. Анизийский ярус), карнийского яруса (См. Карнийский ярус), норийского яруса (См. Норийский ярус) и рэтского яруса (См. Рэтский ярус) (Ладинский ярус выделен в Южных Альпах), отличается очень сложной структурой и чрезвычайным фациальным разнообразием, что затрудняет установление истинной последовательности напластований и палеонтологических зон. В 1960-х гг. в качестве "стандарта для триасового времени" была предложена последовательность палеонтологических зон Северной Америки, где разрезы триасовых отложений более полные, а строение проще.
Табл. 1. - Сопоставление альпийского и среднеевропейского (германского) триаса
Выделенные в Альпах ярусы среднего и верхнего триаса получили всемирное признание; на табл. 2 показано сопоставление зон наиболее изученного верхнего триаса Европы (Альп), Азии (С.-В. СССР) и Северной Америки. Общепризнанных ярусов нижнего триаса нет до сих пор. В Альпах выделяются один (верфенский, или скифский) или два (сейсский и кампильский) яруса; в СССР - индский и оленёкский, в Северной Америке - гризбахский, динерский, смитский и спэтский.
Табл. 2. - Схема сопоставления вернего триаса Европы, Азии и Северной Америки.
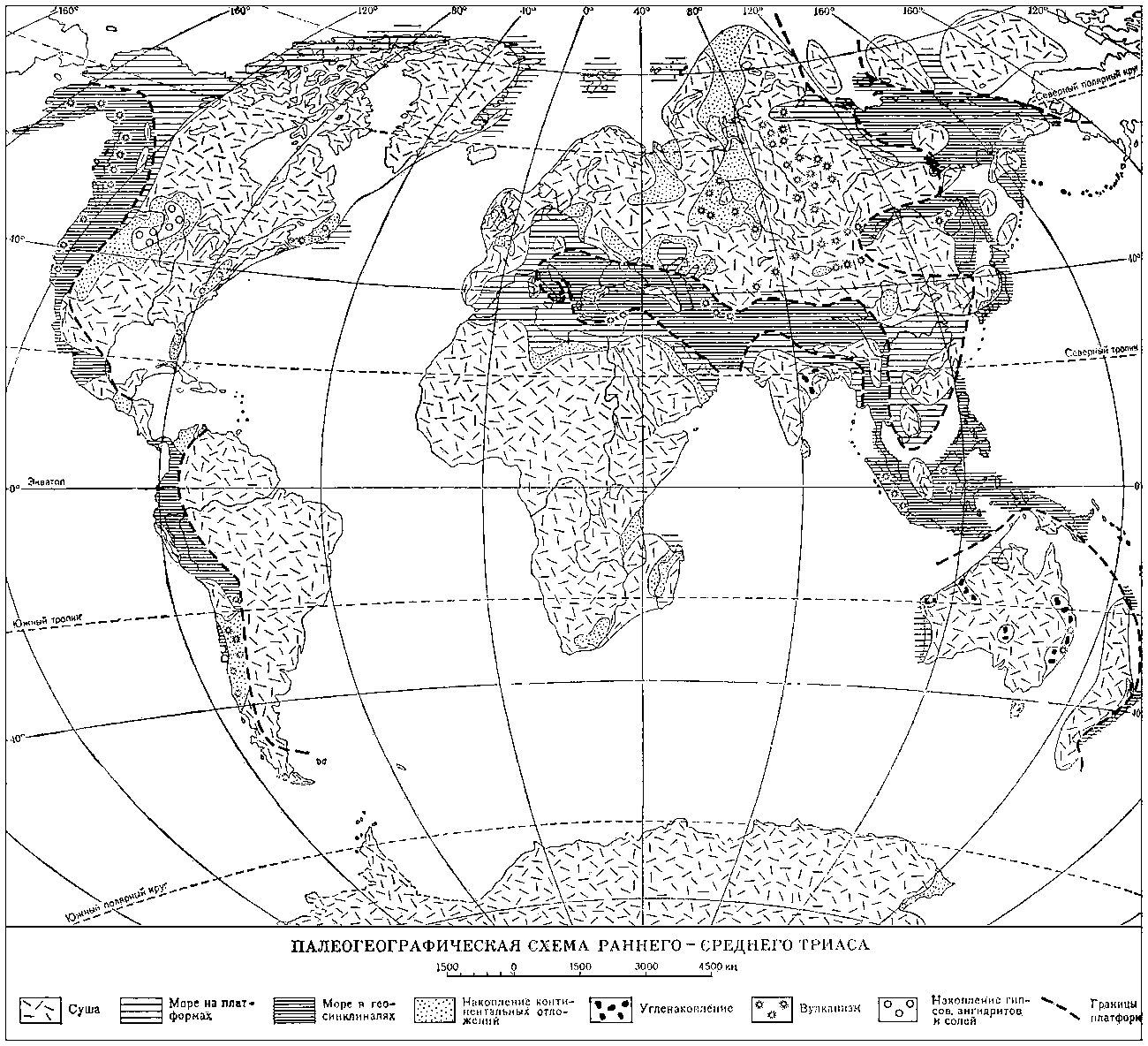
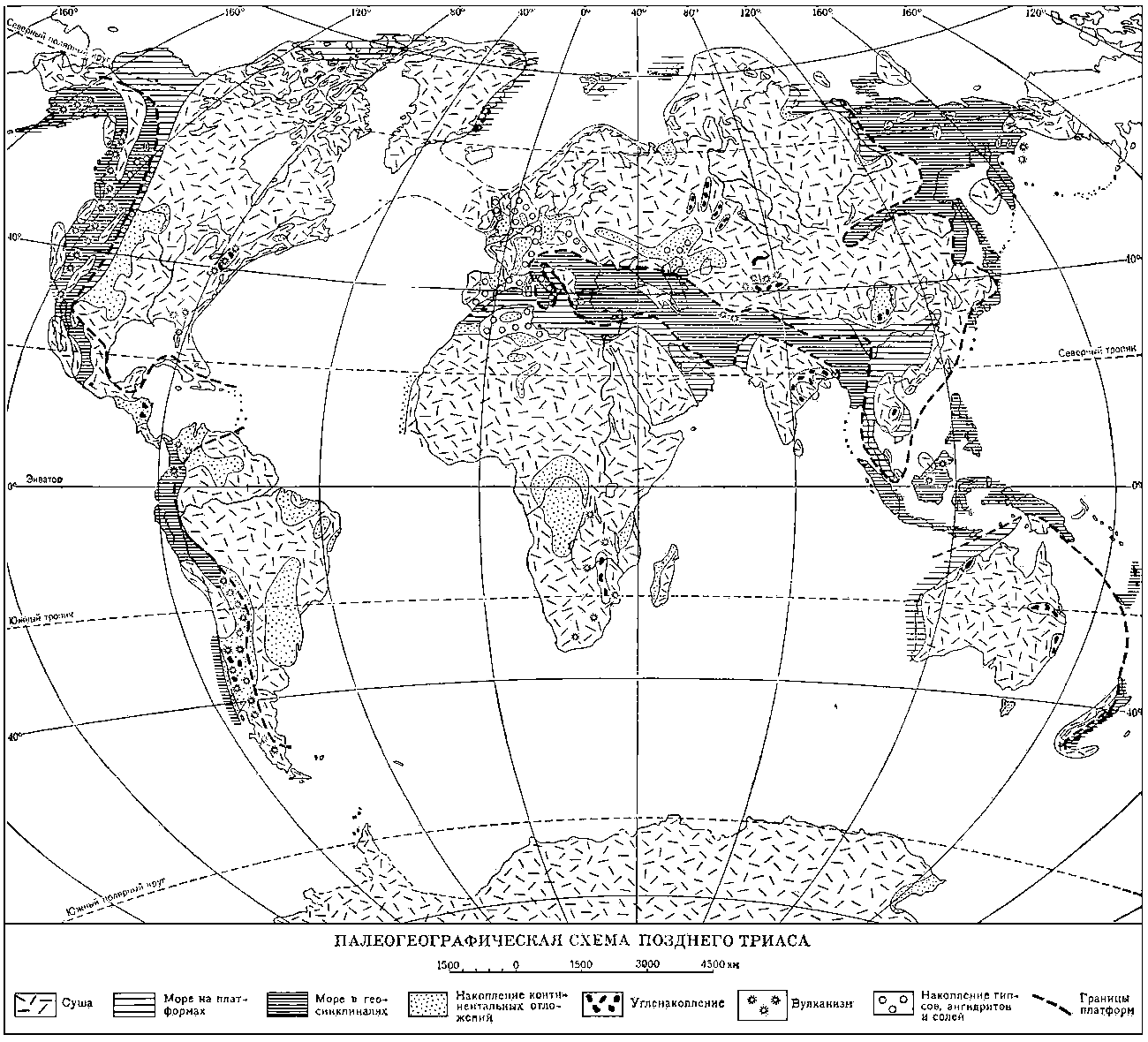
Общая характеристика. К началу триаса герцинские геосинклинальные структуры Западной Европы, Азии (Урало-Тянь-Шаньские и Монголо-Охотские пояса), Северной Америки и Восточной Австралии превратились в молодые платформы; они спаяли в одно целое Лавразию (См. Лавразия) в Северном полушарии, а в Южном полушарии присоединились к Гондване (См. Гондвана). Сильно суженный Средиземноморский геосинклинальный пояс разделял эти две платформы, а Кордильерская и Восточно-Азиатская геосинклинальные области обрамляли их с запада и востока. В триасе континентальные площади, примерно совпадавшие с платформами, господствовали над морскими бассейнами, которые за небольшими исключениями соответствовали геосинклиналям. Морской бассейн Тетис занимал территорию Средиземноморского геосинклинального пояса, временами несколько выходя за его пределы, а в раннем триасе даже не захватывая его полностью. Бореальный бассейн занимал северные части Кордильерской и Восточно-Азиатской геосинклиналей и платформенные области Арктических островов; Тихоокеанский бассейн соответствовал большей части Кордильерской и Восточно-Азиатской геосинклиналей. По берегам Атлантического океана морские триасовые отложения отсутствуют. Это позволяет предполагать, что данного океана в то время ещё не было. Присутствие морских отложений на В. Африки и Мадагаскаре свидетельствует о начале формирования Индийского океана.
Орогенические движения в триасе проявлялись слабо, они имели лишь местное значение. Древнекиммерийский орогенез в конце Т.п. создал складчатые сооружения преимущественно на В. и Ю.-В. Азии.
Герциниды в триасе представляли собой горы, в результате размыва которых на платформах, в отдельных изолированных впадинах накапливались континентальные отложения (пролювиальные, речные, озёрные, лагунные, эоловые). В 1-й половине Т. п. континентальные отложения формировались также в западной части Средиземноморского пояса и восточной части Кордильерской геосинклинали. Песчано-глинистые толщи накапливались в Средней Европе, на Восточно-Европейской платформе, в межгорных впадинах Урала, Северной и Южной Сибири, Северного Казахстана и Средней Азии, где в этих толщах значительна роль эффузивов и туфов. На Сибирской платформе продолжалось формирование трапповой формации, в Восточной Австралии - угленосных отложений, а в Южной Африке и Индии - пёстроцветных толщ. Морские отложения в пределах платформ известны на С. Сибирской платформы, Арктических островах, в Средней Европе, Прикаспии, Восточной Африке и на Мадагаскаре.
Во 2-й половине Т.п. в Средней Европе формировались гипсоносные толщи; угленакопление происходило в Предуралье, на Восточном Урале, на С. и Ю. Сибири, в Средней Азии, на В. Индокитая, на Ю. Аппалачей, в Южной Америке, Южной Африке, Восточной Австралии и Индии, а в Аппалачах, кроме того, проявляется вулканическая деятельность. Морские терригенные отложения известны на Канадском Арктическом архипелаге и Свальбарде.
Карбонатное осадконакопление господствовало в Средиземноморском геосинклинальном поясе, где большое распространение имели рифовые фации, образованные известьвыделяющими водорослями, шестилучевыми кораллами, толстораковинными моллюсками, иглокожими и др. донными организмами. В более глубоких частях моря формировались красноцветные цефалоподовые известняки. В Кордильерской геосинклинали отлагались терригенные и карбонатные осадки, в Восточно-Азиатской геосинклинали - мощные толщи глинистых сланцев, песчаников и конгломератов; карбонатные и кремнистые породы играли подчинённую роль; в верхнем триасе на территории Советского Приморья и Японии накапливались континентальные отложения с углями. В среднем и позднем триасе мощное развитие основного вулканизма характерно для Кордильерской геосинклинали и герцинид Аппалачей и в меньшей степени для Восточно-Азиатской геосинклинали; в Средиземноморском геосинклинальном поясе вулканизм проявлялся слабо.
Климат в Т. п. был достаточно жарким, в 1-й половине - более сухим, во 2-й - более влажным; тропический пояс совпадал с Тетисом и югом Северной Америки, достигая значительной ширины (до 60° к С. и Ю. от экватора).
Органический мир. Во 2-й половине перми началось вымирание палеозойских групп растений и животных, продолжавшееся до конца Т. п. В конце перми и особенно в начале Т. п. появились новые (мезозойские) виды растений и животных, которые в течение Т. п. сосуществовали с древними формами и только с конца Т. п. или начала юры стали господствующими.
Растительный мир суши 1-й половины Т. п. во многом близок к верхнепермскому: исчезли господствовавшие в палеозое древовидные Плауновидные и Каламитовые, Кордаитовые, Прапапоротники и большая часть древних хвойных. Облик растительности в связи с этим изменился коренным образом, хотя новые группы растений ещё не появились: в конце перми и начале триаса преобладали древние хвойные и птеридоспермы (семенные папоротники). Основные мезофитные группы растений (диптериевые папоротники, Саговники, Беннеттиты, Гинкговые, мезофитные хвойные) в значительном количестве появляются во 2-й половине Т. п., но вплоть до конца Т. п. ещё велико значение древних групп (семенных папоротников, древних папоротников и хвойных) и тех групп, расцвет которых приходится на Т.п. (два семейства семенных папоротников, хвощовые). В морях Т.п. большую роль играли рифообразующие водоросли (Доломитовые Альпы).
Для животного мира Т.п. характерен расцвет пресмыкающихся, приспособившихся к жизни на суше и в море, в пресноводных водоемах и в воздухе; некоторые группы земноводных также перешли к жизни в море. Наряду с лабиринтодонтами (См. Лабиринтодонты), котилозаврами (См. Котилозавры), существовавшими ещё в пермское время, появились характерные для мезозоя Архозавры, Ихтиозавры, ряд групп синантозавров, лепидозавров, черепахи. В конце Т. п. появились настоящие костистые рыбы и первые млекопитающие. Пресноводные водоёмы были населены также мелкими ракообразными - ракушковыми (См. Ракушковые) (остракодами), филлоподами.
В морях Т. п. среди беспозвоночных доминировали цератиты (головоногие моллюски), появившиеся в конце перми и вымершие в конце Т. п.; наряду с ними обычны пелециподы, гастроподы, наутилоидеи; в небольшом количестве существовали настоящие аммониты и белемниты; значительно по сравнению с палеозоем уменьшилась роль брахиопод, морских лилий, фораминифер; четырёхлучевые кораллы сменились шестилучевыми.
Биогеографическое районирование. Для триасовых морских бассейнов Тетиса (от Альп до Индонезии) и Ю. Северной Америки характерна обильная и разнообразная фауна беспозвоночных наряду с широким развитием карбонатных рифовых фаций (тропическая область). Другая биогеографическая область (с умеренным климатом) располагалась на территории Бореального бассейна и бассейнов Новой Зеландии и Новой Каледонии, где тропические формы беспозвоночных и рифовые фации отсутствовали. Различие между этими областями четко выступает с конца раннего триаса; граница между ними в течение Т. п. несколько смещалась к С. в Западном полушарии и к Ю. - в Восточном. Флора 1-й половины Т. п. была однообразной.
Начиная с середины Т. п. тропическая флора известна вдоль северного берега Тетиса, а также в Аппалачах и на плато Колорадо. Более умеренная флора заселяла территорию между Тетисом и Бореальным бассейном, а также материки Южного полушария, включая Антарктиду.
Отложения Т. с. в СССР. Морские отложения Т. п. известны по южной и восточной окраинам, а также на С.-В. СССР. В Карпатах развиты главным образом известняки и мергели, в Крыму - песчано-сланцевые отложения; на Северном Кавказе нижняя и верхняя части триаса сложены известняками, средняя-терригенными породами; в Закавказье преимущественно карбонатные толщи, достигающие мощности более 1000 м. Морские терригенные отложения с подчинённой ролью карбонатов известны на Мангышлаке (до 6000 м) и Туаркыре. На Памире в нижней половине разреза преобладают карбонатные осадки, в верхней - терригенные. Мощные толщи (до 3500 м) морских терригенных пород развиты на С.-В. СССР, в Забайкалье и Приморье; в верхах разреза в Приморье чередуются морские и континентальные отложения.
Континентальные отложения Европейской части СССР (центральные районы, Донбасс, Приуралье, Прикаспий) сложены песчано-глинистыми породами с остатками позвоночных, наземных растений, харовых водорослей, остракод, филлопод. В Прикаспии, где мощности Т. с. достигают более 2000 м, в средней части разреза известны морские карбонатные отложения. В изолированных впадинах Восточного Урала, Сибири и Средней Азии нижний триас (до 1400 м) сложен обычно эффузивно-осадочными (в том числе траппы Сибирской платформы и Таймыра), а верхний (до 2700 м) - угленосными отложениями с растительными остатками.
Полезные ископаемые. С отложениями Т. с. связаны месторождения каменных и бурых углей в СССР (Восточный Урал, Южное Приморье), Японии, Вьетнаме, США (Южные Аппалачи), Южной Африке, Восточной Австралии; нефти и газа в СССР (Прикаспий, Дальний Восток), Великобритании, США, Алжире, Ливии; алмазов в СССР (Якутия); урана в США (Колорадо плато); соли в Центральной Европе; медных и медно-никелевых руд в СССР (Норильск), ПНР, США, Канаде; свинцово-цинковых руд в СССР (Дальний Восток), ПНР, Австрии; серебро-свинцовых руд в Югославии; ценных строительных материалов, в том числе знаменитого каррарского мрамора в Италии.
Лит.: Жинью М., Стратиграфическая геология, пер. с франц., М., 1952; Леонов Г. П., Историческая геология, М., 1956: его же. Основы стратиграфии, т. 1, М., 1973; Стратиграфия СССР. Триасовая система, М., 1973; Страхов Н. М., Основы исторической геологии, ч. 2, М.-Л., 1948; Pia J., Grundbegriffe der Stratigraphie, W., 1930; Schmidt M., Die Lebewelt unserer Trias, Öhringen, 1928; Tozer E. Т., A standard for triassic time, Ottawa, 1967; Kozur Н., Probleme der Triasgiiederung und Parallelisierung der germanischen und tethyalen Trias, "Freiberger Forschungshefte", 1974, № 298.
И. А. Добрускина.
Реконструкция ландшафта Восточной Гренландии (скалы Нейл) в рэтском веке: цикадовые, беннеттитовые, гинкговые, папоротники и семенные папоротники.
Скалы Нейл в Восточной Гренландии (современный вид): на переднем плане - замёрзшее море, покрытое снегом.
Реконструкция ландшафта Центральной Европы в раннем триасе: слева на переднем плане - цикадовые, за ними - древовидные плауновидные (плевромеи), дальше - хвойные (вольтция), у водоемов - членистостебельные (схизоневра); справа на переднем плане - следы древнего ящера (хиротерия).
Палеогеографическая схема раннего - среднего триаса.
Палеогеографическая схема позднего триаса.
Руководящие ископаемые триасового периода (1-8 - морские беспозвоночные животные; 9-16 - растения). Головоногие моллюски: 1 - Otoceras из индского яруса Гималаев, уменьшено; 2 - Ceratites из ладинского яруса Центральной Европы, уменьшено; 3 - Tropites из карнийского яруса Альп, уменьшено; 4 - Choristoceras из рэтского яруса Альп, уменьшено. Двустворчатые моллюски: 5 - Daonella из ладинского яруса Альп, уменьшено; 6 - Monotis из норийского яруса Верхоянья, СССР, уменьшено; 7 - Rhaetavicula из рэтского яруса Центральной Европы, уменьшено; 8 - Claraia из верфенского яруса Альп, уменьшено; Водоросли: 9 - Diplopora из ладинского яруса Альп. Плауновидные 10 - Pleuromeia из нижнего триаса Центральной Европы, уменьшено. Папоротники: 11 - Clathropteris из верхнего триаса Вьетнама, уменьшено. Семенные папоротники: 12 - Lepidopteris из верхнего триаса Южной Африки, уменьшено. Беннеттитовые: 13 - Pterophyllum из верхнего триаса Швейцарии, уменьшено. Гинкговые: 14 - Glossophyllum из верхнего триаса Альп, уменьшено; 15 - Sphenobaiera из среднего триаса Южной Ферганы, уменьшено. Хвойные: 16 - Voltzia из нижнего триаса Вогез, уменьшено.
триас
м.
Первый из трех периодов мезозойской эры геологической истории Земли.
Первый из трех периодов мезозойской эры геологической истории Земли.
ТРИАС
а, мн. нет, м. геол.
Первый период мезозоя продолжительностью около 45 млн. лет. Триасовый - относящийся к триасу. | Отложения этого периода состоят из трех компонентов: пестрого песчаника, раковинного известняка и кейпера (толщи континентальных и лагунных отложений); отсюда название.||Ср. ЮРА.
палеонтолог

.")
![Препарация фоссилизированных (ископаемых) костей ''[[Europasaurus]]''](https://commons.wikimedia.org/wiki/Special:FilePath/Europasaurus Praeparation.JPG?width=200 "Препарация фоссилизированных (ископаемых) костей ''[[Europasaurus]]''")

НАУКА О ЖИЗНИ, СУЩЕСТВОВАВШЕЙ ДО НАЧАЛА ГОЛОЦЕНОВОЙ ЭПОХИ ИЛИ В ЕЁ НАЧАЛЕ
Палеоантология; Палеонтолог; Палеонтологи; Палеобиология; Микропалеонтология; Микропалеонтолог
палеоботаник
 (8071723349).jpg?width=200)
. Видны листовые рубцы")
Ботаническая палеонтология; Палеоботаник
м.
Специалист в области палеоботаники.
Специалист в области палеоботаники.
ПАЛЕОНТОЛОГИЯ
НАУКА О ЖИЗНИ, СУЩЕСТВОВАВШЕЙ ДО НАЧАЛА ГОЛОЦЕНОВОЙ ЭПОХИ ИЛИ В ЕЁ НАЧАЛЕ
Палеоантология; Палеонтолог; Палеонтологи; Палеобиология; Микропалеонтология; Микропалеонтолог
биологическая наука, изучающая жизнь прошедших геологических эпох. Отрасль палеонтологии, занимающаяся вымершими животными, называется палеозоологией и может быть разделена на палеонтологию беспозвоночных и палеонтологию позвоночных. Вымершие растения изучает палеоботаника.
Фоссилизация. Окаменелые ископаемые остатки (фоссилии) находят в осадочных горных породах. Чтобы превратиться в окаменелость (фоссилизироваться), организм должен обладать скелетом, раковиной или иными твердыми частями тела и вскоре после смерти быть захороненным, т.е. погрузиться в материал, который будет защищать его труп от разрушительного действия выветривания и эрозии. Мягкие части тела животных сохраняются исключительно редко; например, трупы мамонтов и шерстистых носорогов находят законсервированными в слое вечной мерзлоты в сибирской тундре, где они не разлагаются только благодаря постоянной низкой температуре. К объектам палеонтологии относятся также отпечатки лап и волочащихся частей тела животных, покровов насекомых, перьев и листьев. Некоторые мелкозернистые горные породы, например литографический известняк из Баварии, сохранили отпечатки даже таких нежных структур, как летательные перепонки птерозавров и щупальца древних кальмаров и медуз.
Отливки. Лишь в исключительных случаях ископаемые кости и раковины находят практически в неизмененном виде, обычно же они в то или иной степени разрушаются. После того, как организм занесен осадочным материалом, медленно просачивающаяся вода может растворить и вымыть часть его твердых тканей или даже труп в целом, оставив на их месте полость, напоминающую литейную форму. В дальнейшем какие-либо вещества могут подобно гипсу заполнить ее, образовав естественную отливку. Некоторые отливки весьма точно передают детали строения исходного организма.
Окаменение. Твердые части животных и растений обычно пористые, и грунтовые воды могут заполнить их поры солями кальция, железа или кремнеземом. Такие ископаемые остатки тяжелее и тверже оригиналов. В других случаях грунтовые воды могут растворить весь исходный материал твердых частей организма, оставив на его месте лишь отложенные в порах чужеродные минеральные вещества. Это замещение может быть очень точным и сопровождаться сохранением даже микроскопической структуры объекта. Так дошли до нас некоторые пропитанные кремнеземом стволы деревьев из "окаменелого леса" вблизи Холбрука в Аризоне. Вместе с ископаемыми рыбами, амфибиями и рептилиями иногда находят окаменелые остатки непереваренного содержимого их кишечника, называемые копролитами. Иногда они содержат части скелета съеденных животных.
См. также:
палеонтология
НАУКА О ЖИЗНИ, СУЩЕСТВОВАВШЕЙ ДО НАЧАЛА ГОЛОЦЕНОВОЙ ЭПОХИ ИЛИ В ЕЁ НАЧАЛЕ
Палеоантология; Палеонтолог; Палеонтологи; Палеобиология; Микропалеонтология; Микропалеонтолог
ж.
Научная дисциплина, изучающая растительный и животный мир прошедших геологических эпох.
Научная дисциплина, изучающая растительный и животный мир прошедших геологических эпох.
Палеонтология
НАУКА О ЖИЗНИ, СУЩЕСТВОВАВШЕЙ ДО НАЧАЛА ГОЛОЦЕНОВОЙ ЭПОХИ ИЛИ В ЕЁ НАЧАЛЕ
Палеоантология; Палеонтолог; Палеонтологи; Палеобиология; Микропалеонтология; Микропалеонтолог
(от Палео..., греч. ón, родительный падеж óntos - существо и ...Логия
наука об организмах минувших геологических периодов, сохранившихся в виде ископаемых остатков организмов (См. Ископаемые остатки организмов), следов их жизнедеятельности и Ориктоценозов. Современную П. можно также определить как науку о всех доступных изучению проявлениях жизни в геологическом прошлом на организменном, популяционном и экосистемном (биогеоценотическом) уровнях. В биологии П. предшествует неонтологии (См. Неонтология) - науке о современном органическом мире. По объекту исследования П.- наука биологическая, но возникла она в тесной связи с геологией, широко пользующейся данными П. и вместе с тем служащей главным источником разнообразной информации о среде жизни. Именно эта связь и делает П. целостной наукой о развитии живой природы в геологическом прошлом, без которой невозможно понимание геологической истории биосферы (См. Биосфера), точнее - смены палеобиосфер и становления современной биосферы.
Основные подразделения палеонтологии. В качестве главных подразделений П. выделяют палеозоологию (изучающую ископаемых животных (См. Ископаемые животные)) и палеоботанику (См. Палеоботаника) (посвященную ископаемым растениям (См. Ископаемые растения)). Первая делится на П. беспозвоночных и П. позвоночных; в состав второй входят палеоальгология (ископаемые водоросли), палеопалинология (пыльца и споры древних растений), палеокарпология (семена древних растений) и др. разделы; палеомикология (ископаемые остатки грибов) занимает особое место в системе палеонтологических дисциплин, так как грибы, по мнению многих учёных, образуют самостоятельное царство среди эукариотов (См. Эукариоты). Под условным названием микропалеонтология выделяют раздел П., занимающийся изучением древних микроорганизмов (бентосные простейшие, остракоды, различный зоо- и фитопланктон, бактерии), дисперсных остатков крупных организмов животной и растительной природы и микропроблематик (Конодонты, сколекодонты, Отолиты, хитинозоа и т.п.). Изучение связей организмов прошлого друг с другом и с окружающей средой в рамках популяций, ценозов и всего населения древних бассейнов привело к созданию палеоэкологии. Выявлением закономерностей географического расселения организмов прошлого в зависимости от эволюции климатов, тектоники и др. процессов занимается палеобиогеография. Закономерности захоронения и распространения ископаемых остатков организмов (ориктоценозов) в осадочных толщах изучают Тафономия и биостратономия, следы жизнедеятельности - палеоихнология. Словами с приставкой "палео" часто обозначают разделы систематической П., изучающие остатки древних насекомых (палеоэнтомология), древних моллюсков (палеомалакология), древних рыб (палеоихтиология), древних птиц (палеоорнитология) и т.д. Возможность проникновения в биологическую специфику тканей, морфо-физиологических систем, химизма и т.п. древних организмов привела к появлению палеогистологии, палеофизиологии, палеоневрологии, палеопатологии и др. разделов П. Открытие химической специфики видов и возникновение палеобиохимии позволили подойти к проблемам молекулярной П.
Исторический очерк. Сведения об окаменелостях были известны уже античным философам-натуралистам (Ксенофан, Ксант, Геродот, Теофраст, Аристотель). В эпоху Возрождения, сменившую тысячелетний (5-15 вв.) период застоя, природа окаменелостей получила первую правильную интерпретацию - сперва у китайских натуралистов, а затем и у европейских (Леонардо да Винчи, Джироламо Фракасторо, Бернар Палисси, Агрикола и др.), хотя в большинстве случаев не хватало важнейшего для науки представления о том, что это остатки вымерших организмов. Вероятно, датский натуралист Н. Стено (1669) и английский Р. Гук (опубликован 1705) были одними из первых, кто начал говорить о вымерших видах, а с середины 18 в., с развитием идей М. В. Ломоносова (1763) в России, Ж. Бюффона и Жиро - Сулави во Франции, Дж. Геттона в Великобритании и др., взгляды о постоянных изменениях в живой природе прошлого (теория развития) и значении актуалистического подхода к его познанию, хотя и стихийно, стали завоёвывать всё больше сторонников. Единство системы ископаемых и современных организмов признавал и К. Линней, но он также совершенно отвергал идею изменяемости видов. Решающим периодом для становления П. было начало 19 в., когда У. Смит в Великобритании впервые обосновал определение относительного возраста геологических пластов по окаменелостям беспозвоночных и дал на этой основе первую геологическую карту (1794).
П. как научная дисциплина возникла одновременно и в теснейшей взаимной связи с исторической геологией. Основателем той и другой считают Ж. Кювье, особенно много сделавшего в этих областях в период с 1798 по 1830; в Коллеж де Франс в 1808 он впервые стал читать систематический курс "Истории ископаемых" и на основании глубокого сравнительно-анатомического изучения ископаемых костей млекопитающих фактически создал П. позвоночных. Несколько позднее, с публикацией "Истории ископаемых растений" французского ботаника Адольфа Броньяра, возникла и палеоботаника. Кювье и французский геолог Александр Броньяр (1811) развивали представление о руководящих окаменелостях в геологии; оба они связывали в единой системе ископаемые и современные организмы и оба являлись защитниками гипотезы катастроф (см. Катастроф теория). Термин "П." впервые упомянул (1822) французский зоолог А. Дюкроте де Бленвиль, но распространение он получил лишь после того, как профессор Московского университета Г. И. Фишер фон-Вальдгейм впервые употребил его (1834) вместо термина "петроматогнозия", а во Франции А. Д'Орбиньи начал публикацию сочинений по П. (с 40-х гг. 19 в.).
Создателем первой теории эволюции был Ж. Б. Ламарк, явившийся по существу и основателем П. беспозвоночных. Близким к нему по взглядам был другой эволюционист до-дарвиновского периода Э. Жоффруа Сент-Илер. Однако оба современника Ж. Кювье, также не свободные от известных заблуждений, не могли противостоять его авторитету; в П. 1-й половине 19 в. преобладающей была идея неизменяемости видов и последовательных резких смен в их существовании. Одновременно с накоплением огромного чисто описательного материала в Великобритании, Германии, Франции, Швеции, Италии, России эти общие идеи продолжали энергично развивать швейцарский геолог и палеонтолог Л. Агассис, английский геолог А. Седжвик и особенно французский палеонтолог А. Д'Орбиньи (1840), с именем которого правильнее всего связывать гипотезу катастроф в её завершенном виде (27 переворотов в истории Земли; вывод, основанный на данных о 18 000 видов). Однако положительным результатом этих идей явились формирование стратиграфической П. и завершение разработки уже к началу 40-х гг. общей стратиграфической шкалы Земли. В России успехи П. до-дарвиновского периода связаны с именами Фишера фон-Вальдгейма, Э. И. Эйхвальда, Х. И. Пандера, С. С. Куторги, П. М. Языкова и др. Особое место занимают выдающиеся исследования по стратиграфии, палеонтологии и зоологии предшественника Ч. Дарвина - К. Ф. Рулье, совершенно чуждого идей Креационизма.
П. 60-х гг. 19 в., а затем 20 в. знаменует совершенно новый этап в развитии этой науки. Его начало отмечено появлением наиболее завершенной теории эволюции ("Происхождение видов" Дарвина, 1859), оказавшей огромное влияние на всё дальнейшее развитие естествознания. Хотя многие палеонтологи 19 в., такие, как И. Барранд в Чехии, А. Мильн-Эдвардс и А. Годри во Франции, Р. Оуэн в Великобритании и др., не были дарвинистами, идеи эволюционизма стали быстро распространяться в П. и нашли в ней превосходную почву для своего дальнейшего развития, например в трудах английского естествоиспытателя Т. Гексли, австрийского геолога и палеонтолога М. Неймайра, американского палеонтолога Э. Копа. Но самое выдающееся место, несомненно, принадлежит В. О. Ковалевскому, которого с полным правом называют основателем современной эволюционной П. Только после работ Ковалевского по П. позвоночных и Неймайра по П. беспозвоночных дарвинизм приобрёл ту палеонтологически обоснованную базу, в которой ещё продолжала нуждаться эволюционная теория. Роль П. позвоночных оказалась особенно значительной в разработке теоретических проблем эволюции в связи со сложностью строения не только ныне живущих позвоночных, но и их ископаемых предков. На основе теории эволюции сделаны важные палеонтологические обобщения последователями Ковалевского: бельгийским палеонтологом Л. Долло, американским - Г. Осборном, немецким - О. Абелем и др. В дальнейшем эволюционную палеозоологию в России, а затем в СССР развивали А. П. Карпинский, С. Н. Никитин, А. П. Павлов, Н. И. Андрусов, М. В. Павлова, П. П. Сушкин, А. А. Борисяк, Н. Н. Яковлев, Ю. А. Орлов, Л. С. Берг, А. П. Быстров, И. А. Ефремов, Д. В. Обручев, Л. Ш. Давиташвили, Д. М. Раузер-Черноусова и многие др.; палеоботанику - И. В. Палибин, А. Н. Криштофович, М. Д. Залесский и др. Значительную роль в развитии П. сыграли работы русских биологов А. Н. Северцова, И. И. Шмальгаузена, В. Н. Беклемишева, Д. М. Федотова и др.
Фундаментальной сводкой результатов палеонтологических исследований 19 в. были труды К. Циттеля "Руководство" (1876-1893) и "Основы палеонтологии" (1895). Многократно переиздававшееся последнее издание в полной переработке советских палеонтологов (редактор А. Н. Рябинин) вышло в 1934 на русском языке (беспозвоночные). Наиболее значительным, полностью законченным современным справочным изданием по П. являются "Основы палеонтологии" (15 тт., 1958-64) под редакцией Ю. А. Орлова (Ленинская премия, 1967). Аналогичный 8-томный труд по палеозоологии под редакцией Ж. Пивто издан (1952-1966) во Франции; 24-томное издание по беспозвоночным начало публиковаться в США (с 1953) под редакцией Р. Мура и пока не завершено; переиздаётся с 1970 под редакцией К. Тейхерта.
Основные направления развития палеонтологии и её связи с другими науками. Как наука биологическая П. теснейшим образом связана с комплексом биологических дисциплин (популяционная генетика, биология развития, цитология, биохимия, биометрия и др.), методы которых она частично использует. Всё больше начинают применяться при палеонтологических исследованиях новейшие методики, основанные на использовании различных излучений, химического анализа, электронной и сканирующей микроскопии и др. Традиционны тесные связи и взаимное обогащение со сравнительной анатомией, морфологией и систематикой животных и растений. Морфо-функциональный анализ и изучение морфогенеза скелетных структур ископаемых приводят ко всё более тесным связям П. с физиологией, эмбриологией, биомеханикой. Сравнительно-историческое изучение древних организмов, требующее использования метода актуализма, ведёт ко всё более широким связям П. с экологией, биогеоценологией, биогеографией, гидробиологией и океанологией. Изучение жизни древних морей и современного Мирового океана позволило обнаружить ряд архаических организмов - "живых ископаемых" - латимерия, неопилина, погонофоры и др. Наиболее значительной остаётся связь П., изучающей закономерности исторического развития организмов как в отдельных филумах (генетических рядах организмов), так и в последовательности экологических систем, с эволюционным учением. Филогенез и экогенез в одинаковой мере не могут быть достаточно поняты без объединения достижений П. и неонтологии. История филогенетических построений, начиная от первой чисто неонтологической схемы Э. Геккеля (1866) и до современных частных и общих построений филогении, показывает, сколь шаткими оказываются эти схемы без достаточных палеонтологических знаний. Вместе с тем для самой П. важно правильное понимание таких явлений, как параллелизм в изменчивости (см. Гомологических рядов закон), парафилия, внутривидовой полиморфизм и т.д., имеющих то или иное значение в формировании представлений о происхождении и родословной биологических таксонов. П. и неонтологию тесно объединяют общие и важнейшие в биологии проблемы видообразования, факторов и темпов эволюции, её направлений. Однако можно с уверенностью сказать, что П. получила от неонтологии значительно больше, чем неонтология от неё пока взяла и могла бы взять. П. обладает совершенно неисчерпаемым фондом фактических документов действия эволюционного процесса (только ископаемых беспозвоночных известно не менее 100 тыс. видов), и неонтология (даже сравнительная анатомия и систематика) ещё далека от освоения этого фонда. Неонтологией явно недостаточно оценена фактическая длительность эволюционного процесса, а она теперь прослеживается документально почти с границы химической и биологической эволюции на протяжении 3,5 млрд. лет; история прокариотов, эукариотов и становления многоклеточных организмов. (Metaphyta и Metazoa) фиксируется в П. уже датами изотопной геохронологии. Наконец, сама система и родословные отношения органического мира не могут оставаться без существенной перестройки в свете палеонтологической истории организмов дофанерозоя и фанерозоя. Многие проблемы неонтологии не возникли бы без П. (темпы и направления эволюции, происхождение высших таксонов органического мира).
Значение П. в системе наук о Земле не менее велико. Геология стала подлинно исторической наукой о Земле только с возникновением стратиграфии (См. Стратиграфия) на рубеже 18 и 19 вв., когда был найден способ определения относительной хронологии геологических образований по ископаемым остаткам организмов (Руководящие ископаемые) и возникла объективная возможность геологического картирования не типов горных пород по их петрографическим признакам, а возрастных подразделений слоистой оболочки земной коры. Стратиграфическая корреляция, по данным П. и вспомогательным данным изотопной хронометрии и др. физических методов сопоставления древних отложений, лежит в основе успехов геологии. Коренное значение для внедрения П. в стратиграфическую геологию имело эволюционное учение, опиравшееся на теорию естественного отбора, концепцию необратимости эволюционного процесса; сама геология такой теории не имела. Французский палеонтолог и геолог А. Оппель, изучавший юрские отложения Центральной Европы, впервые предложил зональный Палеонтологический метод сопоставления отложений, и, хотя зональная стратиграфия не получила быстрого распространения на всю стратиграфическую шкалу, эта идея П. стала ведущей во всём дальнейшем совершенствовании общей стратиграфической шкалы и в региональной стратиграфической корреляции. Отсюда берёт своё начало научная Биостратиграфия, хотя сам термин был предложен бельгийским палеонтологом Долло лишь в 1909. П. внесла в геологию свой метод отсчёта времени (биохронология), и современная так называемая хроностратиграфическая шкала, строго говоря, является шкалой биостратиграфической. Палеонтологический метод оказался наиболее универсальным как для обоснования самих стратиграфических подразделений и выявления коррелятивных особенностей их биологической характеристики (периодичность или этапность развития органического мира), так и для конкретной типизации (стандартизации) биостратиграфических границ, что стало важнейшей международной задачей стратиграфии. Экологический контроль оказывает всё возрастающее влияние на палеонтологический метод в региональной стратиграфии, а биогеографический - на межрегиональную и планетарную корреляцию отложений. При этом выявляется теснейшая связь П. с учением об осадочных фациях (само определение последних невозможно без данных П.), с литологией и седиментологией вообще, геохимией и биогеохимией осадочных пород. Данные П. играют важнейшую роль во всех палеогеографических реконструкциях, в том числе и палеоклиматологических (выявление сезонности и климатической зональности по данным скелетных структур животных, палеодендрологии, географии древних организмов и т.п.). Литолого-фациальные карты, наряду с их огромным значением в исторической геологии, становятся всё более важными и для прогноза поисково-разведочных работ на уголь, нефть, газ, бокситы, соли, фосфориты и др. полезные ископаемые. При этом остаётся важной породообразующая роль самих древних организмов (многие типы карбонатных и кремнистых пород, залежи различных каустобиолитов (См. Каустобиолиты), проявление фосфатности и различной минерализации, связанной либо прямо с первичным физиологическим химизмом древних организмов, либо с последующими адсорбционными процессами в органогенных скоплениях). Органический мир древних эпох и его непосредственное участие в ведущих процессах биосферы создали главный энергетический потенциал Земли. Связь П. с геологией нерасторжима не только потому, что последняя является главным поставщиком палеонтологического материала и фактической информации об условиях среды обитания в различные периоды (а без этого невозможно развитие П., так же как и неонтологии), но и потому, что геология пока остаётся и главным потребителем результатов палеонтологических исследований, ставя перед ними всё более новые и сложные задачи, требующие освоения современной биологии и геологической теории.
Научные учреждения и общества. Имеется большое количество палеонтологических обществ: Палеонтографическое общество в Великобритании (создано в 1847; с 1957 Палеонтологическая ассоциация), Швейцарское палеонтологическое общество (1874), секция П. в Венском зоолого-ботаническом обществе (1907), секция П. в Геологическом обществе США (1908; с 1931 Общество прикладной П. и минералогии и отдельно Палеонтологическое общество), Палеонтологическое общество Германии (1912), Русское (ныне Всесоюзное) палеонтологическое общество (1916), Палеонтологическое общество Китая (1929) и т.д. Большую роль играет Московское общество испытателей природы (с 1940 имеется палеонтологическая секция). Такие общества имеются почти во всех развитых и в ряде развивающихся стран. С 1933 они связаны с единой Международной палеонтологической ассоциацией (МПА), деятельность которой особенно активизировалась после Генеральных ассамблей (они всегда проходят вместе с сессиями Международных геологических конгрессов) в Нью-Дели (1964), Праге (1968), Монреале (1972). МПА связана с Международными союзами геологических и биологических наук. Она имеет большое количество корпоративных членов и специализированные международные исследовательские группы (на базе соответствующих комиссий и комитетов), которые становятся главной формой международной деятельности МПА (симпозиумы, конференции и пр.), поддерживаемой национальными палеонтологическими (как в Чехословакии, Польше и др. странах) или геологическими (как в СССР) комитетами и университетами. МПА объединяет научные интересы свыше 6000 палеонтологов, из них около 40\% советских. Советское отделение МПА входит в её состав на правах континентального отделения, и его президент является вице-президентом ассоциации.
Научные исследования в области П. ведутся главным образом в учреждениях национальных геологических служб и компаний, геологических и биологических институтах академий наук, а также в горно-геологических вузах и музеях (например, палеонтологические отделы Британского музея, Американского естественноисторического музея в Нью-Йорке, Смитсоновского института Естественноисторического музея в Вашингтоне, Народного музея в Праге, Зенкенбергского музея во Франкфурте-на-Майне, Естественноисторического музея в Будапеште, Палеонтологического музея в Осло, Музея Онтарио в Торонто; в СССР - Музей им. Ф. Н. Чернышева Центрального научно-исследовательского Геологоразведочного института в Ленинграде, Палеонтологический музей Зоологического института АН УССР в Киеве и др.). Большую роль играют палеонтологические отделы и лаборатории многих университетов мира: Калифорнийского, Канзасского, Мичиганского и др. в США; Аделаидского, Канберрского, Сиднейского в Австралии; Лундского, Стокгольмского в Швеции, а также Токийского, Мадридского, Витватерсрандского в ЮАР, Ла-Плата в Аргентине и многих др.; в СССР - Московского, Ленинградского, Киевского, Томского и т.д. Имеются самостоятельные специализированные палеонтологические институты: Палеонтологический институт АН СССР, институт палеобиологии АН Грузинской ССР, Палеонтологический институт в Бонне (ФРГ), институт палеонтологии человека в Париже и институт палеонтологии Естественноисторического музея Франции, Палеоботанический институт Индии, институт палеозоологии Польской АН, Палеобиологический институт в Упсале (Швеция), институт палеонтологии позвоночных и палеоантропологии и Геолого-палеонтологический институт в КНР, палеонтологические институты в университетах Вены, Милана, Модены, в Университете им. Гумбольдта в Берлине, институты геологии и палеонтологии в ряде университетов ФРГ (Гёттингене, Тюбингене, Киле, Штутгарте, Марбурге, Мюнстере) и в др. странах.
Планомерные палеонтологические исследования в России начались с созданием Геологического комитета в Петербурге (1882) и учреждением при нём с 1912 штатных должностей палеонтологов (Н. Н. Яковлев, М. Д. Залесский, А. А. Борисяк и др.), хотя уже в Кунсткамере Петра I стали сосредоточиваться остатки "допотопных животных". В 1917 в Геологическом комитете впервые в стране была создана крупная палеонтологическая секция. Вместе с Русским палеонтологическим обществом (1916), Горным институтом, первой в России университетской кафедрой П. Петроградского университета, организованной в 1919 М. Э. Янишевским, и остеологическим отделом Геологического и минералогического музея АН секция стала главным центром распространения работ по П. и самоопределения П. в дочерних учреждениях Геологического комитета (Всесоюзный научно-исследовательский геологоразведочный институт и др.), а также в АН СССР. В 1930 А. А. Борисяком был создан в Ленинграде первый специальный Палеозоологический (современное название - Палеонтологический) институт, наиболее полно развернувший свои исследовательские и экспедиционные работы после переезда АН в Москву и привлечения к работе московских палеонтологов. Однако основной рост палеонтологических лабораторий, секций, отделов и кадров шёл в геологических учреждениях министерства геологии СССР, АН СССР и союзных республик, различных ведомств и на геологических факультетах университетов. Крупнейшее значение имело создание сети различных микропалеонтологических лабораторий (первой - в Нефтяном геологоразведочном институте, ныне Всесоюзный научно-исследовательский геологоразведочный институт в Ленинграде, в 1930), отделов палеонтологии и биостратиграфии в Геологическом институте АН СССР (Москва), институте геологии и геофизики Сибирского отделения АН СССР (Новосибирск), институте геологии АН Эстонской ССР (Таллин), институте геологии АН Казахской ССР (Алма-Ата) и многочисленных аналогичных подразделений в др. центральных и региональных учреждениях АН и Геологической службы СССР, а также в учреждениях биологических (Ботанический институт АН, Ленинград. Институты биологического профиля Дальневосточного научного центра АН, Владивосток, и др.) и географических (Институт географии АН, институт океанологии АН, Москва, и др.). Палеонтологи СССР работают более чем в 200 учреждениях, около 90\% из них связаны с науками о Земле. В научной и координационной деятельности в П. основное значение имеют ежегодные тематические сессии Всесоюзного палеонтологического общества в Ленинграде, собирающие до 600 участников, и Научный совет Отделения общей биологии АН по проблеме "Пути и закономерности исторического развития животных и растительных организмов", объединяющий все специализированные палеонтологические комиссии и проводящий свои пленарные сессии раз в пять лет в Москве, а также ВСЕГЕИ, координирующий в течение многих лет работу территориальных геологических управлений.
Периодическая печать. Важнейшими специальными изданиями по П. являются в СССР: "Палеонтологический журнал" (с 1959), "Ежегодник Всесоюзного палеонтологического общества" (с 1917) и "Труды" его годичных сессий (с 1957), "Палеонтология СССР" (с 1935), монографические серии по П. многих институтов; за рубежом: "Acta palaeontologica polonica" (Warsz., с 1956), "Palaeontologia Polonica" (Warsz., с 1929); "Acta palaeontologica sinica" (Peking, с 1962), "Vertebrata Palasiatica" (Peking, с 1957), "Palaeontologia Sinica" (Peking, с 1922), "Rozpravy. Ústředniho ústavu geologickeho" (Praha, с 1927), "Annales de palé́ontologie" (P., с 1906), "Revue de micropaléontologie" (P., с 1958), "Bulletins of American Paleontology" (lthaca - N. Y., с 1895), "Journal of Paleontology" (Tulsa, с 1927), "Micropaleontology" (N. Y., с 1955), "Palaeontographica Americana" (lthaca, с 1916), "Palaeontographical Society Monographs" (L., с 1847), "Palaeontology" (Oxf., с 1957),"Palaeobiologica" (W., 1928-45), "Palaeogeography, palaeoclimatology, palaeoecology" (Amst., с 1965), "Palaeontographia italica" (Pisa, с 1895), "Rivista italiana di paleontologia e stratigrafia" (Mil., с 1895), "Palaeontologische Abhandlungen" (В., с 1965), "Palaeontographica" (Stuttg., с 1846), "Palaeontologische Zeitschrift" (Stuttg., с 1914), "Senckenbergiana Lethaea" (Fr./M., с 1919), "Biomineralisation" (Stuttg.- N. Y., с 1970), "Palaeontologia indica" (Delhi, с 1957), "Journal of Palaeontological Society of India" (Lucknow, с 1956), "Lethaia" (N. Y.- L., с 1968), "Palaeontologia mexicana" (Mex., с 1954), "Palaeontologia africana" (Johannesburg, с 1963), "Paleontological Bulletins" (Wellington, с 1913), "Ameghiniana" (Buenos Aires, с 1957) и др. Не меньшее количество работ по П. публикуется в изданиях общего характера по геологии, зоологии и ботанике. Современный уровень исследований по П. хорошо отражают "Proceedings of the International Paleontological Union" (Warsz., с 1972), "International Geological Congress Sect. Paleontology" (Montreal, 1972) и труды других национальных или международных съездов палеонтологов в СССР, США, Франции, Великобритании и др. странах. Ведётся постоянный раздел "Палеонтология" в реферативном журнале Всесоюзного научно-исследовательского института технической информации (1954-73).
Лит.: История. Борисяк А. А., В. О. Ковалевский. Его жизнь и научные труды, Л., 1928; Давиташвили Л. Ш., История эволюционной палеонтологии от Дарвина до наших дней, М.- Л., 1948; Криштофович А. Н., История палеоботаники в СССР, М., 1956; Павлов А. П., Полвека в истории науки об ископаемых организмах, М., 1897; Zittel К., Geschichte der Geologic und Paläontologie bis Ende des XIX Jahrhunderts, Münch.- Lpz., 1899.
Руководства. Друщиц В. В., Обручева О. П., Палеонтология, 2 изд., М., 1971; Методика палеонтологических исследований, пер. с англ., М., 1973; Основы палеонтологии. Справочник для палеонтологов и геологов СССР, [т.] 1-15, М., 1958-64; Палеонтология беспозвоночных, М., 1962; Glaessner М. F., Principles of micropalaeontology, N. Y.- L., 1963; Müller A. H., Lehrbuch der Paläozoologie, Bd 1-3, Jena, 1957-70; OIson E. C., Vertebrate paleozoology, N. Y.- L.- Sydney, 1971; Raup D. М., Stanley S. M., Principles of paleontology, S. F., 1971; Traite de paleontologie, publ. sous la dir. de J. Riveteau, t. 1-7, P., 1952-69; Treatise on invertebrate paleontology, ed. R. C. Moore, Lawrence (Kansas), 1953-69, ed. C. Teichert, 2 ed., Lawrence (Kansas), 1970-72.
Общие работы. Борисяк А. А., Основные проблемы эволюционной палеонтологии, М.- Л., 1947; Давиташвили Л. Ш., Причины вымирания организмов, М., 1969; Красилов В. А., Палеоэкология наземных растений, Владивосток, 1972; Палеонтология, М., 1972; Палеопалинология, т. 1-3, Л., 1966; Современные проблемы палеонтологии, М., 1971; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М.- Л., 1964; Шмальгаузен И. И., Происхождение наземных позвоночных, М., 1964; Atlas of palaeobiogeography, ed. A. Hallam, Amst., 1973; Brooks J. and Shaw G., Origin and development of living systems, L.- N. Y., 1973; Evolution and environment, ed. E. T. Drake, New Haven - L., 1968; Floristics and paleofloristics of Asia and Eastern North America, ed. A. Graham, Amst., 1972; Kuźnicki L., Urbanek A., Zasady nauki о ewolucji, t. 1-2, Warsz., 1967-70; Lehman J.-P., Les preuves paleontologiques de l'évolution, P., 1973; Organisms and continents through times, L., 1973; Proceedings of the North American paleontological convention, ed. E. L. Yochelson, v. 1-2, Lawrence (Kansas), 1970-71; Termier H., Termier G., Biologie et écologie des premieres fossiles. P., 1968.
Палеоэкология и тафономия. Вялов О. С., Следы жизнедеятельности организмов и их палеонтологическое значение, К, 1966; Геккер Р. Ф., Введение в палеоэкологию, М., 1957; Ефремов И. А., Тафономия и геологическая летопись, кн. 1, М.- Л., 1950; Организм и среда в геологическом прошлом, отв. ред. Р. Ф. Геккер, М., 1966; Среда и жизнь в геологическом прошлом, Новосиб., 1973; Яковлев Н. Н., Организм и среда, 2 изд., М.- Л., 1964; Ager D. V., Principles of paleoecology, N. Y.- L., 1963; Reyment R. A., Introduction to quantitative paleoecology, Amst.- [a. o.], 1971; Schäfer W., Aktuo-Paläontologie nach Studien in der Nordsee, Fr./M., 1972; Trace fossils, ed. T. P. Crimes, J. C. Harrer, Liverpool, 1971.
Микропалеонтология. Вопросы микропалеонтологии, в. 1-16, М., 1956-73; Fichier micropaleontologique general, P., 1943-71; Pokorný V., Grundzüge der zoologischen Micropaläontologie, Bd 1-2, B., 1958; Proceedings of the First International conference on planktonic microfossils, v. 1-2, Leiden, 1969.
Справочники, библиография. Коробков И. А., Палеонтологические описания, 2 изд., Л., 1971; Маир Э., Принципы зоологической систематики, пер. с англ., М., 1971; Палеонтологи Советского Союза. Справочник, сост. И. Е. Занина, Л., 1968; Палеонтологический словарь, М., 1965; Бжеленко Л. К., Митрошина Л. Н., Шевырев А. А., Палеозоология СССР. Библиография отечественной литературы за 1917-1967 гг., кн. 1-2, М., 1971-1973; Lehmann U., Paläontologisches Wörterbuch, Stuttg., 1964: Directory of palaeontologists of the World-1972, lerusalem, 1973.
Б. С. Соколов.
Википедия
Триасовый период

Триа́совый пери́од (триа́с) — первый геологический период мезозойской эры. Следует за пермским периодом и предшествует юрскому. Начался 251,902±0,024 млн лет назад, закончился 201,3±0,2 млн лет назад. Продолжался, таким образом, около 51 млн лет. Совокупность отложений (горных пород) триасового возраста называется триа́совой систе́мой.
Введён Фридрихом Альберти в 1834 году. Назван по наличию трёх слоёв в континентальных отложениях этого времени в Западной Европе: пёстрого песчаника, раковинного известняка и кейпера.

